First discovered in Western Australia in the mid-1880s, the bizarre-toothed eugeneodont cartilaginous fish Helicoprion davisii was initially mistaken for a species of the equally weird Edestus. It was eventually recognized as part of a separate genus over a decade later, when similar fossils of its close relative Helicoprion bessonowi were found in the Ural Mountains.
1890s-2000s
With Helicoprion only known from strange buzzsaw-like spiral whorls, the function and location of this structure in the fish’s body was a huge source of confusion for over a century.
The earliest interpretation was a defensive structure curling upwards from the snout, then as part of the tail or dorsal fins. It was soon realized to probably be part of the lower jaw instead, but the exact arrangement was still a mystery.
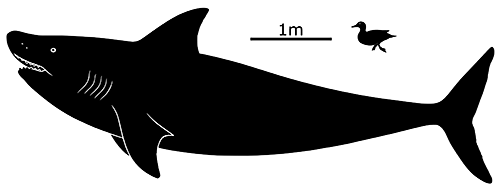
A downward-curling position was popular in reconstructions for much of the 20th century. From the 1960s onwards, however, discoveries of preserved skull cartilage and soft-tissue body outlines of other eugeneodont species began to give a better idea of what these fish were and what they looked like. They were identified as being related to modern chimaeras, but with a very different appearance – they had streamlined shark-like bodies with large triangular pectoral fins, a single dorsal fin, no pelvic or anal fins at all, and broad keels along the sides of their tails.
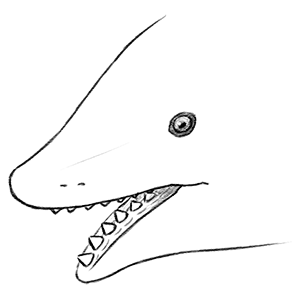
A single tooth whorl sat in the middle of the lower jaw, with its sides covered by skin, and the largest and youngest teeth formed at the back before spiralling forwards, downwards, and inwards.
In the 1990s a “pizza-cutter” reconstruction gave Helicoprion long narrow jaws with the whorl positioned very far forwards, sawing and crushing prey against the underside of the snout. A version with the whorl very deep inside the throat was also proposed in 2008, but only a year later a new variant of the pizza-cutter saw-jaw model suggested the presence of a specialized “pocket” in the upper jaw lined with teeth.
2020s
Finally, in 2013, CT scanning of a Helicoprion specimen originally discovered in the 1960s revealed something incredibly special – an almost complete three-dimensionally preserved and articulated set of jaws. It showed narrow jaws that were shorter than the previous reconstructions, with the whorl occupying the entire lower jaw and braced by cartilage on each side.
We now know Helicoprion davisii was found worldwide during the early-to-mid Permian, about 272-268 million years ago, and based on some of the biggest known tooth whorls it may have reached sizes of up to 8m long (~26′), similar in size to modern basking sharks. It continuously added new and larger teeth to its whorl throughout its life, with the smaller older teeth being retained instead of shed, slowly pushed into a tight spiral deep inside the lower jaw.
The upper jaw formed a sheath-like pocket lined with a “pavement” of numerous tiny rounded teeth, and as Helicoprion closed its jaws the various parts of the whorl simultaneously grabbed, sliced, and pulled prey further into its mouth – a mechanism possibly specialized for efficiently de-shelling cephalopods like ammonites and nautiloids.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}