Probably around 2m long (~6’6″), it had a short deep skull and massive cheekbones – unusual features for an early saurischian, and convergently similar to those of some later theropods.
Along with its close relatives Tawa, Chindesaurus, and Daemonosaurus (a grouping collectively known as “morphoraptorans”) it represents one of the latest-surviving known herrerasaurians, suggesting that these early-diverging carnivorous dinosaurs persisted in low-latitude regions until much closer to the end of the Triassic than previously thought.
Despite its very mole-like appearance, this little mammal was a member of an extinct lineage known as palaeanodonts — and its closest living relatives are actually pangolins.
Around 15cm long (~6″), Xenocranium was highly adapted for a subterranean burrowing lifestyle, with an upturned shovel-shaped snout bearing a pad of thickened skin, and short powerful limbs with large digging claws. Its eyes were very reduced, functionally blind, and may not have even been visible in life. Its sense of hearing was also specialized for the sort of low-frequency sounds that carry well through the ground.
It was probably a head-lift digger, using upward motions of its snout and downward strokes with its forelimbs to excavate tunnels while foraging for worms and underground insects.
Ubaghsicystis segurae was an echinoderm that lived during the mid Cambrian, about 506-497 million years ago, in what is now Spain. Similar fossils are also known from Morocco, Mexico and Canada, suggesting this genus was quite widespread in marine continental shelf habitats at the time.
It was part of an extinct group known as eocrinoids, which were the earliest known echinoderms to develop stalked bodies and specialized feeding appendages — but despite the name and similar body plan, their resemblance to crinoids was due to convergent evolution rather than any direct relation.
Its globular body was only about 5mm in diameter (~0.2″), with a long stalk at least twice that length ending in a small disc that attached it to the substrate. The upper half of its body was dotted with small holes (called epispires) that probably served a respiratory function, and unlike most other eocrinoids it seems to have had just two slender flexible feeding appendages.
It would have been a suspension feeder, using tube feet to catch food particles floating around in the water, then move it down to its mouth positioned at the base of the two “arms”.
Living in what is now North America during the Late Triassic, about 237-216 million years ago, Poposaurus grew to around 4.5m long (~15′) with roughly half of that length taken up by just its long tail. With its sharp-toothed jaws, small arms, bipedal locomotion, and counterbalancing tail, it convergently evolved the same sort of body plan and ecology as carnivorous theropods – which were still in their early days at the time, and wouldn’t really become the dominant terrestrial predators until after the end-Triassic extinction.
Unlike most other pseudosuchians Poposaurus lacked bony osteoderm armor, seems to have been capable of a digitigrade posture, and its claws were flattened and somewhat hoof-like, all adaptations that suggest it was built for running after fast-moving prey.
Like other therizinosaurids it would have been a chunky-bodied herbivore with a small beaked head atop a long neck, long rake-like claws on its hands, stout legs, and a rather short tail. But it was rather small compared to most of its close relatives, estimated at about 3m long (~9’10”), with its known fossil remains including several vertebrae, partial ribs and pelvis, and a set of nearly-complete arms and hands.
Its hands had only two well-developed fingers, with a small splint-like vestigial third finger, an anatomical condition convergently seen in some other theropod groups but previously unknown in therizinosaurids. One of its long curved claws also preserved a rare example of a thick keratinous sheath, showing that in life the claw was over 40% longer than its bony core.
Duonychus’ elbow and finger joints had a fairly limited range of motion – more similar to the forearms of Tyrannosaurus than other therizinosaurids – but its claws were able to flex almost 90° at the tips of its fingers, which may have given it the ability to reach out and grab onto foliage with a very strong and precise grip.
Although the cartilaginous skeletons of chondrichthyans rarely preserve, the exceptional preservation conditions of the Bear Gulch Limestone fossil deposits mean we do actually have full-body soft tissue impressions of this species. It was about 1m long (3’3″) with an unusually slender eel-like body, a pointed snout, no dorsal fins, and an elongated tapering tail.
Preserved gut contents show that Thrinacodus gracia preyed on shrimp-like crustaceans and smaller cartilaginous fish such as Falcatus and Harpagofututor. It would have inhabited a shallow tropical bay environment, and may have had a similar sort of lifestyle to the modern eels it resembled, hiding in crevices or burrowing into sediment and ambushing passing prey.
About 13cm long (~5″), it had a tall narrow disc-shaped body convergently similar to modern reef fish, and it was mostly scaleless with only a few scales on its underside, below its pectoral fins, and along the top of its tail. It also appears to have been toothless, and probably used its large scissor-like jaws to snip off mouthfuls of soft food such as algae.
But its most distinctive feature was its highly elongated pennant-like dorsal fin, which may be an example of sexual dimorphism – fossils of short-finned individuals have also been found, and although they were originally named as a separate species (Dorypterus althausi) they probably actually represent female D. hoffmani.
Up to about 15cm long (~6″), its tapering tadpole-like body plan somewhat resembled that of modern knifefishes and featherbacks, with the top part of its tail fin highly elongated into a ribbon-like shape and the rest of its tail fins being vestigial. The distinctive humped shape of its back was also much more pronounced in larger, more mature individuals.
It was probably a fairly slow swimmer, and preserved gut contents suggest it mainly ate small soft-bodied prey.
Its closest known relative seems to have been the eel-like Holopterygius – but since around 60 million years and different continents separated them both, this suggests the existence of a whole ghost lineage of other tapering coelacanths yet to be discovered.
Apatemyids were a group of unique early placental mammals that lived during the first half of the Cenozoic, known from North America, Europe, and Asia. Due to their specialized anatomy their evolutionary relationships are rather murky (they were traditionally part of the convoluted mess that was “Insectivora”), but currently they’re thought to be a very early offshoot of the Euarchontoglires, the branch of placentals that includes modern rodents, lagomorphs, treeshrews, colugos, and primates.
Living in what is now western Europe during the mid-Eocene, around 47 million years ago, Heterohyus nanus was a small apatemyid about 30cm long (~12″) – although just over half of that length was made up of its tail.
Like other apatemyids it had a proportionally big boxy head, with large forward-pointing rodent-like incisors in its lower jaw and hooked “can-opener-shaped” incisors in its upper jaw.
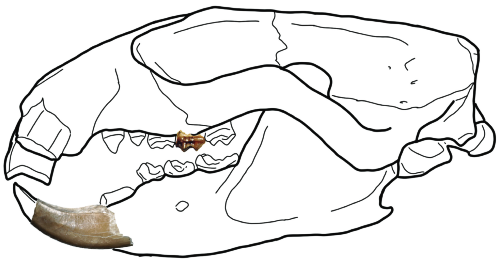
Example of an apatemyid skull from the closely related American genus Sinclairella. From Samuels, Joshua X. “The first records of Sinclairella (Apatemyidae) from the Pacific Northwest, USA.” PaleoBios 38.1 (2021). https://doi.org/10.5070/P9381053299
The rest of its body was rather slender, and fossils with soft tissue preservation from the Messel Pit in Germany show that it had a bushy tuft of longer fur at the end of its long tail.
But the most distinctive feature of apatemyids like Heterohyus were their fingers, with highly elongated second and third digits resembling those of modern striped possums and aye-ayes. This suggests they had a similar sort of woodpecker-like ecological role, climbing around in trees using their teeth to tear into bark and expose wood-boring insect holes, then probing around with their long fingers to extract their prey.
It was around 2m long (6’6″) and, despite its very tapir-like appearance and lack of horns, it was actually closer related to modern rhinoceroses – it was part of a group of early rhino-cousins known as amynodontids, which convergently evolved both hippo-like and tapir-like lifestyles.
Cadurcodon was the most tapir-like of the tapir-like amynodontids, with a short deep skull and retracted nasal bones that indicate it had a well-developed prehensile trunk. Males also had large tusks formed from their upper and lower canine teeth, which may have been used for fighting each other.