

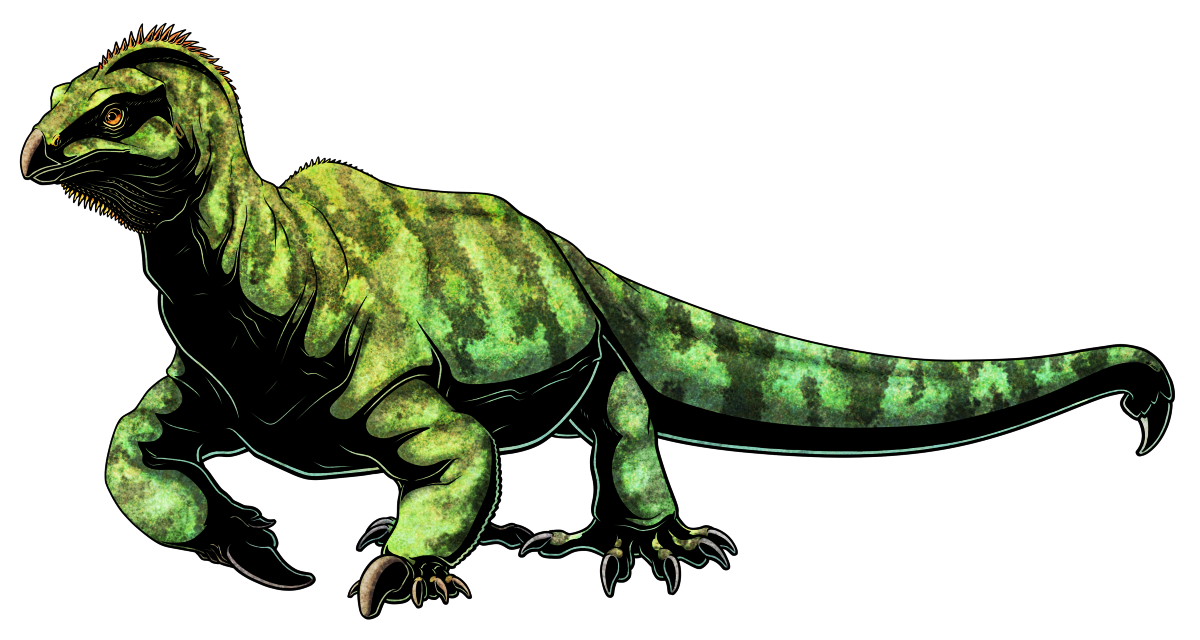
Akidostropheus oligos was a small tanystropheid archosauromorph reptile that lived during the late Triassic, about 223-218 million years ago, in what is now Arizona, USA.
Only a few tiny isolated vertebrae have been discovered, so its full size and appearance isn’t known – making any reconstruction rather speculative – but it was probably around 30cm long (~12″). Like other tanystropheids it would have been a long-necked lizard-like animal, and may have had a similar build to the closely-related Tanytrachelos.
But despite the scarcity of material the few known vertebrae are unique among archosauromorphs, bearing elongated spikes with a surface texture that suggests they were covered with keratinous sheaths. The spikes were conical, sharp, and hooked on the neck and upper back, but became more flattened, straighter, and blade-like on the lower back and tail.
These structures were probably defensive in nature, especially considering that there’s direct fossil evidence for predators targeting the long necks of tanystropheids and decaptiating them.
Akidostropheus lived in a tropical floodplain environment around a meandering river system, but without more and better fossils it’s impossible to tell what its ecology was. Tanystropheids were a strange and diverse bunch, with both terrestrial and aquatic lifestyles, bipedal runners, and possibly even bizarre leg-gliders, so this spiky little Triassic weirdo could have been doing almost anything.
Continue reading “Akidostropheus”










{kind=link}
{kind=link}