It was around 2m long (6’6″) and, despite its very tapir-like appearance and lack of horns, it was actually closer related to modern rhinoceroses – it was part of a group of early rhino-cousins known as amynodontids, which convergently evolved both hippo-like and tapir-like lifestyles.
Cadurcodon was the most tapir-like of the tapir-like amynodontids, with a short deep skull and retracted nasal bones that indicate it had a well-developed prehensile trunk. Males also had large tusks formed from their upper and lower canine teeth, which may have been used for fighting each other.
Originally represented by three statues, there are two surviving originals of the Eocene-aged palaeotheres depicting Plagiolophus minor (the smaller sitting one) and Palaeotherium medium (the larger standing one).
The sitting palaeothere unfortunately lost its head sometime in the late 20th century, and the image above shows it with a modern fiberglass replacement. Then around 2014/2015 the new head was knocked off again, and has not yet been reattached – partly due to a recent discovery that it wasn’t actually accurate to the sculpture’s original design. Instead there are plans to eventually restore it with a much more faithful head.
These early odd-toed ungulates were already known from near-complete skeletons in the 1850s, and are depicted here as tapir-like animals with short trunks based on the scientific opinion of the time. We now think their heads would have looked more horse-like, without trunks, but otherwise they’re not too far off modern reconstructions.
This sculpture went missing sometime after the 1950s, and its existence was almost completely forgotten until archive images of it were discovered a few years ago. Funds were raised to create a replica as accurate to the original as possible, and in summer 2023 (just a month before the date of my visit) this larger palaeothere species finally rejoined its companions in the park.
Compared to the other palaeotheres this one is weird, though. Much chonkier, wrinkly, and with big eyes and an almost cartoonish tubular trunk. It seems to have taken a lot of anatomical inspiration from animals like rhinos and elephants, since in the mid-1800s odd-toed ungulates were grouped together with “pachyderms“.
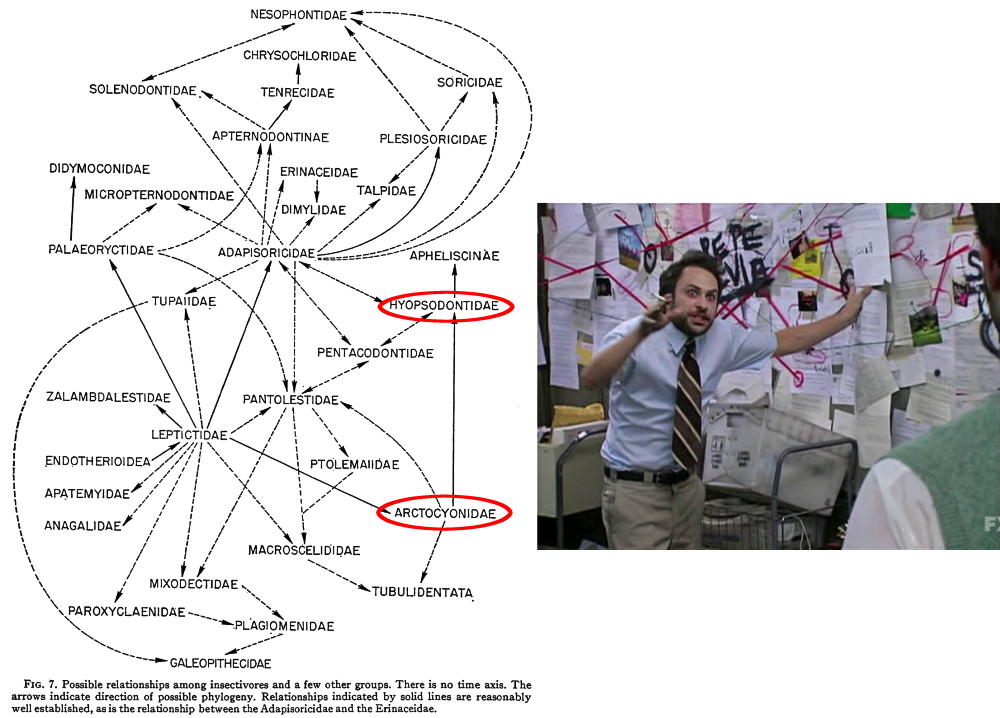
“Insectivora” was a wastebasket taxon so bad it had to be revised multiple times, but there’s another particularly infamous case in mammal taxonomy that’s still in the process of being resolved – the “condylarths“.
This group was first created in the early 1880s, during the Bone Wars, and initially was just a subgroup of odd-toed ungulates containing only the phenacodontids. But just a few years later Condylarthra was promoted up to its own order, and groups like the periptychids and hyopsodontids were added in too.
Then over the next few decades various groups were added and removed from the condylarths, most notably with the mesonychids and arctocyonids being brought in from their previous position with the creodonts.
By the mid-20th century the condylarths had become a big convenient dumping ground for any and all “primitive” ungulate-like mammals that didn’t easily fit into any modern groups, ranging in age from the early Paleocene through to the early Oligocene. But it soon became apparent that they had the same problem as the “insectivores” – there weren’t really any unique anatomical features that united all these animals together.
They generally had rounded-cusped molar teeth and hoof-like toes, but they also had rather generalized “primitive mammal” features and a diverse range of ecologies. Some were small herbivores, but others were coati-like or dog-like omnivores, and some were even bear-sized carnivores.
From left to right, top row: Hyopsodus lepidus (hyopsodontid), Meniscotherium chamense (phenacodontid), Arctocyon primaevus (“arctocyonid”). Bottom row: Ectoconus ditrigonus (periptychid), Mesonyx obtusidens (mesonychid)
It wasn’t even clear how the various different condylarth groups were actually related to each other. The best guess was that arctocyonids had arisen from within the “insectivores”, with a Protungulatum-like form as the common ancestor of all the other condylarths. Where exactly modern ungulates had then evolved from within the condylarths was also still uncertain.
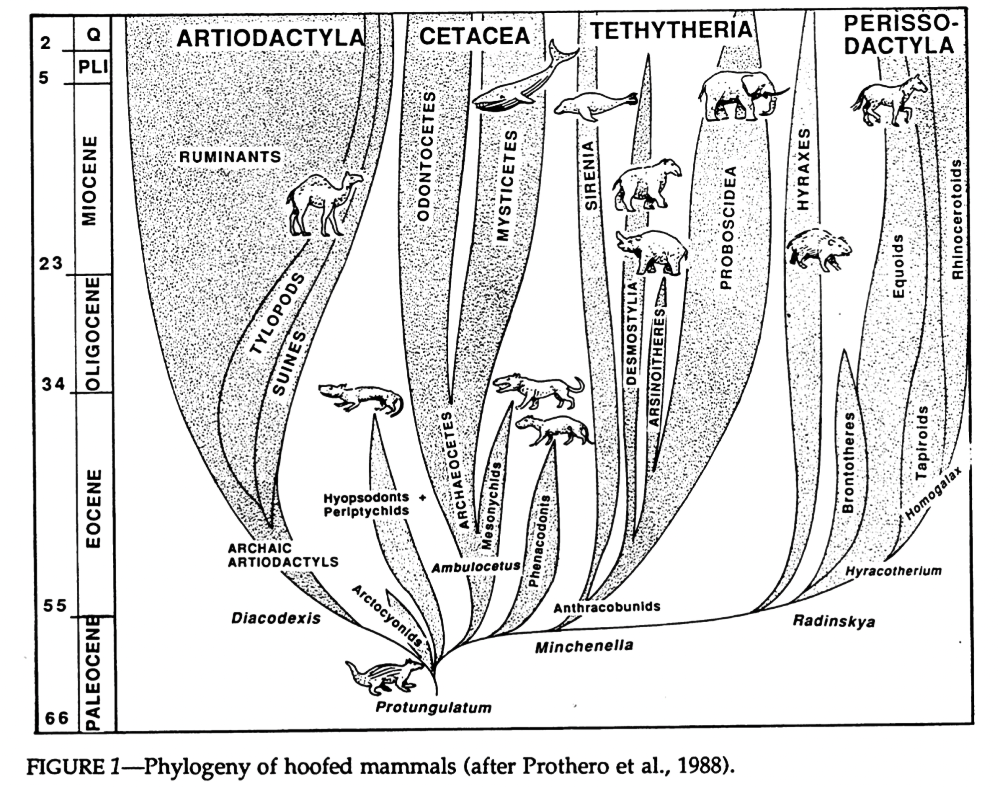
Cladistic analysis in the 1980s began to tackle the confusing pile of assorted condylarths, and showed that they weren’t the single ancestral source of all modern ungulates, but instead a loose collection of several unrelated groups from all over the ungulate evolutionary tree. Arctocyonids, periptychids, and hyopsodontids were placed as early “primitive” lineages, phenacodontids were loosely linked with the ancestors of odd-toed ungulates once again, and mesonychids were considered to be the ancestors of whales.
And, once again paralleling the mess of the “insectivores”, it wasn’t until genetic methods became available in the late 1990s that larger-scale ungulate relationships began to be properly resolved. The paenungulates (elephants, hyraxes, and sirenians), which had been traditionally considered to be a major branch of ungulates, were removed entirely and reclassified as afrotheres. And, along with some new fossil discoveries, whales were recognized as having actually evolved from within the even-toed ungulates instead of from mesonychids.
This shake-up threw the still-problematic “condylarth” classifications back into question – with some “condylarths” turning out toalso be afrotheres instead of true ungulates.
Today the actual relationships of the main “condylarth” ungulate families are still in the process of being figured out, and there’s a lot of remaining uncertainty and disagreement about them.
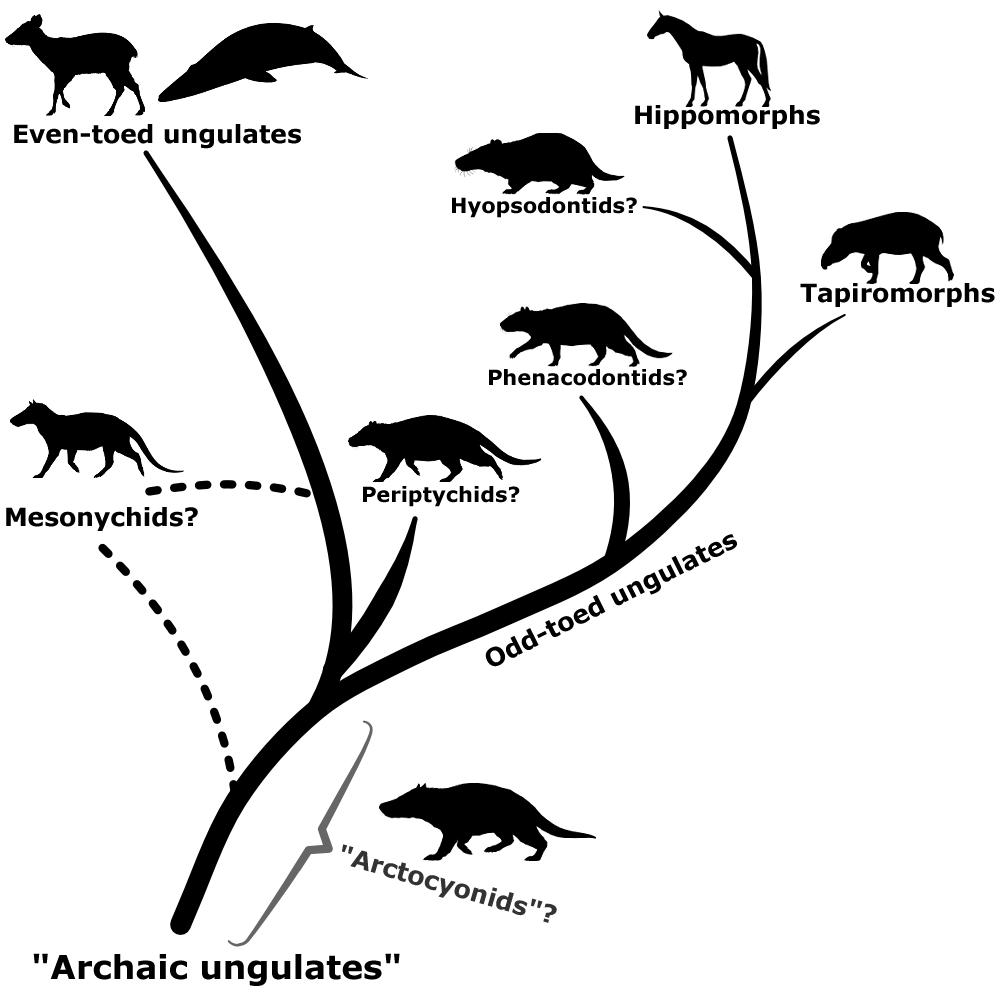
Phenacodontids seem to have mostly maintained their traditional position as early odd-toed ungulates, and hyopsodontids may potentially be part of this group too – possibly as members of the hippomorph lineage, closely related to horses and brontotheres. Arctocyonids might be a wastebasket themselves, with some studies finding them to be a mix of several different archaic ungulate lineages. Periptychids may have links to the even-toed ungulates. The mesonychids, meanwhile, are now generally considered to be a separate order from the traditional “condylarths”, and may be either an early branch of the even-toed ungulates or much more basal ungulates closely related to the “arctocyonids”.
Since the term “condylarth” no longer has any real taxonomic meaning some paleontologists have proposed replacing it with “archaic ungulate” to distance from the historical messiness of the old name. But this hasn’t really caught on, and many papers still use “condylarth” in a very loose sense to refer to an “evolutionary grade” of early ungulates of unclear evolutionary affinities.
And while that’s the last main entry for this month, we’re not quite done yet. There’s still one weekday left in October, and after digging through so many taxonomic garbage cans there’s only one place we can go now.
Squeezing in one last bonus #Spectember post this year!
This one isn’t based on a specific prompt, but instead is a companion piece to a previous one.
While North American brontotheres were adapting to the spread of grasslands, some of their Asian cousins took a very different evolutionary path through the rest of the Cenozoic.
Today’s #Spectember concept is a combination of a couple of anonymous submissions:
Crowned brontothere (left) and woolly paracerathere (right)
These two animals are the descendants of brontotheres and paraceratheres, almost the last living representatives of their kinds, hanging on in the equivalent of modern-day times in a world similar to our own.
Elasmotherium sibiricum was a giant rhinoceros that lived during the mid-to-late Pleistocene epoch, between about 800,000 and 39,000 years ago. Found across much of the Eurasian steppe dry grassland environments, it stood around 2.5m tall (8’2″) at the top of its humped shoulders and weighed about 4 tonnes (4.4 US tons), making it close in size and mass to a modern elephant.
It was the last known representative of a particularly ancient lineage of rhinos, last sharing a common ancestor with modern forms over 40 million years ago.
A large bony dome on its forehead is traditionally thought to have supported an enormous keratinous horn like the distantly-related woolly rhino, but a 2021 study has recently challenged that interpretation. The dome structure was actually rather thin-walled and wouldn’t have been able to support the weight of a giant horn, instead probably being covered by a much stumpier backwards-pointing nub – while an enlarged nasal cavity inside the dome also suggests it may have actually functioned as a resonating chamber, similar to the crests of hadrosaurs or the extinct wildebeest Rusingoryx.
It also had a smaller toughened “pad” on its nose that may have been used along with a prehensile upper lip to dig around in the soil for plant roots and tubers.
This particular genus was very widespread for much of the Eocene, found across Europe, Asia, and North America, crossing back and forth between the continents via the North Atlantic land bridge.
The Jamaican Hyrachyus lived during the mid-Eocene, around 45 million years ago, and was very anatomically similar to the North American Hyrachyus affinis – with the known fossil material not being considered distinct enough to be assigned to a new species yet. It was also about 15-20% smaller than its mainland relative, standing only 25cm tall at the shoulder (10″), but it’s not yet clear if this was a case of insular dwarfism or not.
Its presence in ancient Jamaica suggests that there may have been some sort of land connection between the proto-island and Central America during the early Eocene, when a chunk of what would eventually become western Jamaica was located much closer to the coasts of Honduras and Nicaragua. It’s the only fossil ungulate known from the Caribbean, and one of only a few terrestrial mammals in the region with North American evolutionary roots (the others being the extinct rodents Caribeomys merzeraudi and Oryzomys antillarum, and modern solenodons).
Unfortunately these little rhinos didn’t get much time on their island home. Jamaica subsided fully underwater about 40 million years ago, drowning its unique Eocene ecosystem entirely, and wouldn’t re-emerge and be re-colonized until much later in the Cenozoic.
While the largest animal known to ever exist is an aquatic mammal (the modern blue whale), mammals on land have never managed to attain the same sort of massive sizes seen in the sauropod dinosaurs. This is probably due to a combination of factors, including their reproductive strategies, metabolisms, and physiological differences like lacking internal air sacs – but even being limited to overall smaller body sizes, some of the mega-mammals known to have evolved during the Cenozoic were still absolutely enormous.
(The exact name of this animal has a long and complicated history, and in various times and places it’s also been known as Indricotherium, Baluchitherium, and Pristinotherium.)
Found across much of Eurasia during the Oligocene, about 34-23 million years ago, Paraceratherium was part of an ancient lineage of long-legged hornless rhinoceroses. It stood around 4.8m tall at the shoulder (15’9″) – big enough that most modern humans would be able to walk right underneath its belly without even having to duck – and it had elongated limbs and a long neck that gave it an overall appearance much more like a giant weird horse than a rhino.
There was a pair of downward-pointing tusks at the front of its upper jaw, and the shape of the nasal region of its skull suggests its nose formed a short prehensile tapir-like trunk, which would have been used to help grab and strip leaves from high branches.
I’ve also reconstructed it here with a speculative dewlap on its neck, used for both display and thermoregulation.
Back during the early Eocene, around 50 million years ago, global temperatures were much warmer than today, and in North America tropical and subtropical rainforests extended as far as Alaska.
And one of the most abundant animals in these balmy ecosystems was a small mammal called Hyopsodus, an early type of ungulate that was probably part of the perissodactyl lineage, closely related to the ancestors of modern horses.
Many different species of this genus have been discovered, ranging from rat-sized to cat-sized. Remains of Hyopsodus account for up to 30% of fossils in some locations, with tens of thousands of specimens known – although most of them are isolated teeth and jaw fragments.
(The illustration here depicts Hyopsodus wortmani, a 30cm/12″ long species which lived about 50-46 million years ago across the Western and Southern USA.)
More substantial skeletal remains of this little mammal are very rare, and initially seemed to show a long weasel-like body that resulted in Hyopsodus being given the nickname of “tube-sheep”. But more recentspecimens have given us a better idea of its proportions, and it wasn’t really tubular at all. Instead it was probably built more like a cavy or a hyrax, with a more chunky body and a spine held more strongly curved.
Its teeth suggest it was a generalist omnivore, probably mainly eating a mixture of vegetation, fruits, seeds, insects, and occasionally smaller animals, and while its limbs were proportionally short it was likely still quite an agile fast-moving animal. It also appears to have had some ability to dig, and may have sheltered in burrows similarly to modern groundhogs.
But one of the most surprising things about the “tube-sheep” comes from studies of its braincase via CT scans of its skull. Its brain was unusually large for its size, and had enlarged areas associated with good senses of smell and hearing – and notably one sound-processing region (known as the inferior colliculus) was developed to a degree similar to those seen in echolocating animals.
Analysis of its ear bones suggest it wasn’t highly specialized for echolocation like bats, but may have still been capable of a more basic shrew-like version, using it for close-range navigation.
Much like Platybelodon from a few entries back, chalicotheres look like a fictional creature design rather than something that actually existed.
These animals were odd-toed ungulates related to modern horses, tapirs, and rhinos, who ranged across Africa, Eurasia, and North America for a large chunk of the Cenozoic. Instead of hooves they had large claws on their feet, and they appear to have occupied the same sort of ecological niche as ground sloths or therizinosaurs – sitting or rearing up on their hind legs to browse on high vegetation, using the hook-like claws on their forelimbs to pull down and strip branches.
Tylocephalonyx skinneri here was one of the latter group, known from the Miocene of North America about 16-13 million years ago. Standing about 2m tall at the shoulder (6’6″), it had the same sort of chunky body as other schizotheriines and walked around with its large front claws held up to keep them raised away from the ground.
But there was also an unusual feature on its otherwise rather horse-like head – a large bony dome on top of its skull, like a mammalian version of a pachycephalosaur.
It probably used its dome in the same way as the dinosaurs it convergently resembled, headbutting or flankbutting in fights with each other.












{kind=link}
{kind=link}