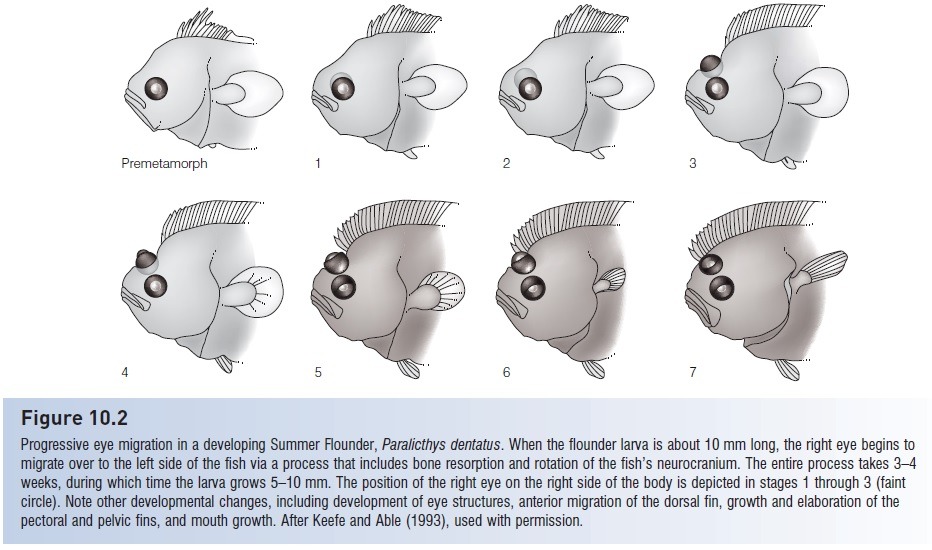
Modern flatfish are characterized by their highly asymmetrical skulls, with both eyes positioned on just one side of their head. They aren’t actually born this way, but instead they undergo “eye migration” as juveniles, twisting up their skulls to bring one eye across the top of the head.
Progressive eye migration in a developing Summer Flounder, Paralicthys dentatus. From Helfman et al (2009). The diversity of fishes. 2nd ed., Wiley-Blackwell.
This bizarre arrangement is the result of flatfish adapting to life laying flat on the seafloor, but instead of slowly widening and flattening themselves out they took an evolutionary “shortcut” by simply tipping their tall narrow bodies over onto one side. Initially this would have left one of their eyes unusable, but random mutations causing slightly asymmetrical skulls would have rapidly become highly advantageous to the earliest members of this lineage – and over time they just got wonkier and wonkier.
We’ve even found fossils of early flatfish in the “halfway there” stage of their evolution!
Amphistium paradoxum lived in what is now northern Italy during the Eocene, around 50-48 million years ago. About 20cm long (~8″), it had one eye partially migrated towards the top of its head, but not all the way around yet, showing a transitional state between its bilaterally symmetric ancestors and its more twisted-skulled modern relatives.
Unlike most modern flatfish Amphistium came in both “right-eyed” and “left-eyed” forms in equal numbers, suggesting that a genetic preference for a specific side also hadn’t developed yet.
But one of the most bizarre of all was the genus Nipponites, whose ribbed shell looked like a bundle of tangled asymmetrical coils.
Nipponites bacchus lived in what is now Hokkaido, Japan, during the late Cretaceous about 90 million years ago. Around 10cm long (~4″), its shell was less tightly coiled up than its better-known relative Nipponites mirabilis, but these looser whorls were formed in the same way via a series of U-bends in different directions during its growth.
Despite their irregular and ungainly appearance, the unique shape of these ammonites seems to have actually been very hydrodynamically stable. They weren’t fast-moving, but they didn’t need to be, probably spending most of their time floating suspended in the water column catching small planktonic prey from around themselves.
The genus Walliserops was one of the weirdest-looking trilobites, covered in numerous pointy spines and sporting a large three-pronged “trident” on the front of its face.
They also had some degree of asymmetry in their bodies. Their tridents often didn’t fork evenly, and their long forehead spines curved off to one side – possibly so they could lift their heads up without stabbing themselves in the back.
Walliserops hammii lived in what is now Morocco during the early-to-mid Devonian, about 403-392 million years ago. Around 5cm long (~2″) It was one of the “short trident” species of Walliserops, and its chunky forehead spine curved particularly strongly to the right.
The function of these trilobites’ elaborate tridents is still poorly understood. But an unusual individual of the long-tridented species Walliserops trifurcatushas been found with a lopsided four-pronged trident, and since it was able to grow to full maturity the shape of the structure probably wasn’t absolutely vital for survival, suggesting it wasn’t used for feeding or sensory purposes.
The tridents may instead have been used for combat with each other similar to the horns of some modern beetles. However, these sorts of features are usually only seen in males, and there’s currently no definite evidence for any significant sexual dimorphism in trilobites.
(Although perhaps like ceratopsid dinosaurs their ornaments were just present in both males and females, being also useful for species recognition, visual display, and defense against predators.)
Represented today by starfish, brittle stars, sea urchins, sea cucumbers, and crinoids, the echinoderms have a characteristic five-way radial symmetry that makes them barely even recognizable as bilaterians. Their true ancestry is only revealed by their genetics and their larvae, which still retain bilateral symmetry – and the way they metamorphose into adults is bizarre, essentially growing a whole new radial body from within the left side of their larval body.
(Sea cucumbers and sand dollars are superficially bilateral as adults, but evolved this secondarily on top of their existing radial symmetry. And some adult echinoderms like starfish also seem to retain a little bit of “behavioral bilaterism”, generally preferring to move with a specific arm always acting as their “front” end.)
The first known echinoderms appeared in the fossil record during the early Cambrian, about 525 million years ago, but the common ancestor of the whole group probably actually originated a few tens of millions of years earlier in the mid-to-late Ediacaran. Early echinoderms seem to have started off as flattened animals that sat on the seafloor filter-feeding, and with this largely immobile way of life their bodies started to shift into asymmetry, no longer constrained by the locomotory advantages of being bilaterally symmetric.
In fact, for these early sedentary filter-feeders being radial was actually much more advantageous, able to distribute sense organs all around their bodies and grab food from any direction without having to reposition themselves, converging on the lifestyle of non-bilaterian cnidarian polyps. The evolutionary transition from bilateral to asymmetrical to pentaradial seems to have happened incredibly quickly during the Cambrian Explosion, and all modern echinoderms probably evolved from a group called the edrioasteroids, maintaining their new base body plan even when they later began taking up more mobile lifestyles again.
But during the process of all that some very alien-looking lineages split off at various stages of anatomical weirdness.
Stylophorans had asymmetrical bodies with a single feeding arm at the front, and varied from irregular boot-like shapes to almost bilateral heart shapes depending on their specific ecologies. The highly asymmetrical forms were probably spreading their weight out over soft soupy mud in quiet waters, while the more bilateral forms may have been more streamlined to deal with stronger water currents.
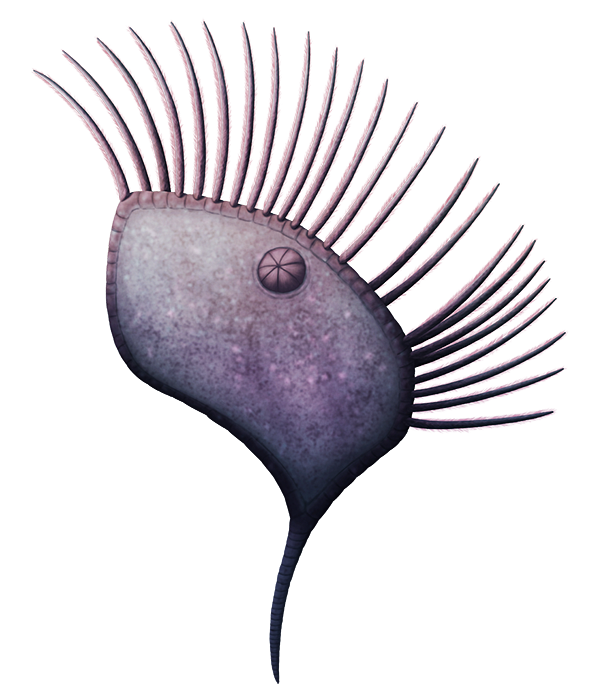
Sokkaejaecystis serrata
Sokkaejaecystis serrata was a stylophoran that lived during the late Cambrian, about 501-488 million years ago, in what is now South Korea. It was tiny, only about 1cm long (~0.4″), and its boot-shaped body was surrounded by spines and flanges that spread out its surface area and probably also made it much more awkward for small predators to attempt to eat.
Meanwhile the solutes started off as immobile animals living attached to the seafloor via a stalk-like appendage. But fairly early in their evolution they switched to a more active mode of life, modifying their stems into tail-like “steles” that were used to push themselves along.
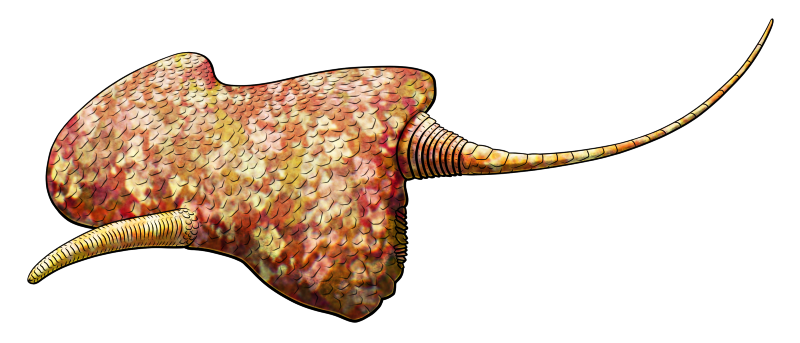
Maennilia estonica
Maennilia estonica lived in what is now Estonia during the late Ordovician, about 450 million years ago. It was quite large for a solute at about 12cm long (~4.7″), with a sort of vaguely-trapezoidal body, a short feeding arm, and a long thin stele.
Both of these strange early echinoderm lineages were surprisingly successful, surviving for a good chunk of the Paleozoic Era alongside their more familiar radial relatives. The solutes lasted until the early Devonian about 400 million years ago, and the stylophorans continued all the way into the late Carboniferous about 310 million years ago.
Probably evolving from Siberion-like lobopodians, the dinocaridids were an “evolutionary grade” of panarthropods that were closely related to the ancestors of true arthropods. These animals were characterized by specialized front appendages on their heads and large swimming lobes along the sides of their segmented bodies, and their group included some of the most famous of the Cambrian “weird wonders”.
The earliest branches of the dinocaridids were the “gilled lobopodians”, which had lobopodian-like legs on their undersides and gills on the upper surfaces of their body lobes. The flap-like structures may have initially evolved just to provide a larger surface area for respiration, but they were quickly co-opted for swimming purposes and opened up a whole new range of ecological opportunities to the ancestral dinocaridids.
And some of the most confusing of them were the paracrinoids, which evolved an incredibly diverse range of body shapes during their group’s relatively short 40 million year existence during the Ordovician.
Despite the name these echinoderms weren’t particularly closely related to true crinoids, instead being part of a completely extinct lineage known as the blastozoans. Their ancestors had been radially symmetric, but paracrinoids largely abandoned that body plan, instead developing irregularly shaped and often asymmetric bodies ranging from round to flattened. They had between two and five “food grooves” on their upper surfaces, derived from the ambulacra, lined with numerous feeding appendages along only the left side of each.
They were shallow-water animals, living either attached to the seafloor by a long stem or anchored into the sediment by a shorter one, suspension feeding with their appendages and transporting the food particles towards the mouth located between the bases of the food grooves.
(…And speaking of mouths, some paracrinoid species appear to have had two of them.)
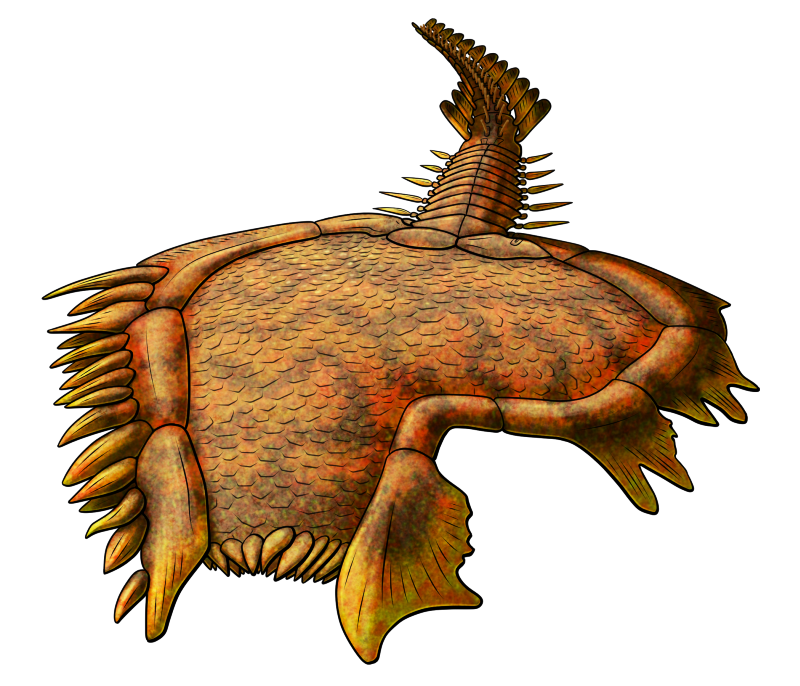
About 10cm tall (4″), it lived on the seafloor in sheltered waters protected from strong waves by large reefs, and is unusual even among its weirdo relatives for features such as retaining feeding appendages on both sides of its food grooves – although irregularly arranged and with fewer on one side than the other. Its body was shaped rather like a flattened bean, with two food grooves diverging from roughly the centre of the top margin, chunky skeletal plates forming a border around its edges, and a short stem at is base.
It also had an unusually large “anal pyramid” on the opposite side of its body from its mouth, and this may have been used for respiration as well as waste expulsion, similar to modern sea cucumbers.
Thylacocephalans were a bizarre group of extinct marine arthropods that often looked like tiny alien creatures – and whose evolutionary relationships are still uncertain. Despite existing in oceans around the world for at least 350 million years, their fossil record is rather spotty and their internal anatomy is often poorly preserved, making it difficult to figure out anything more specific than “probably some sort of crustacean“.
Possible thylacocephalans are known from as far back as the Cambrian, but one of the earliest definite members of the group was Thylacares brandonensis.
Living during the early Silurian, around 436 million years ago, in the region that today is part of Wisconsin, USA (found in the same fossil formation as last week’s Venustulus), this species measured up to about 7.5cm long (3″). Its body was enclosed by a large bivalved carapace, with protruding stalked eyes and what may have been a pair of antennae, along with smaller raptorial limbs than its later relatives.
While it was less specialized than other thylacocephalans it was probably a similar sort of swimming predator, catching prey with its spiny limbs.
For the final entry in this series, let’s take a look at a modern weird-headed species – and where better to find some of the strangest and most unique-looking animals alive today than the deep sea?
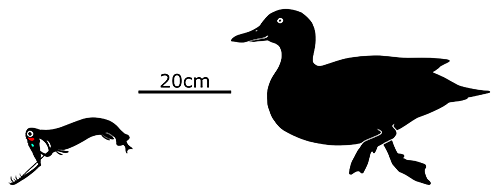
Malacosteus, also known as the stoplight loosejaw, is a 25cm long (10″) genus of dragonfish found at depths of over 500m (1640′) in oceans all around the world, with the exception of the Mediterranean and polar waters. Two different species are currently recognized, with Malacosteus niger here known from just below the Arctic Circle down to the southern reaches of the subtropics, and Malacosteus australis ranging from there to around 45°S, and up towards the equator in the Indian Ocean.
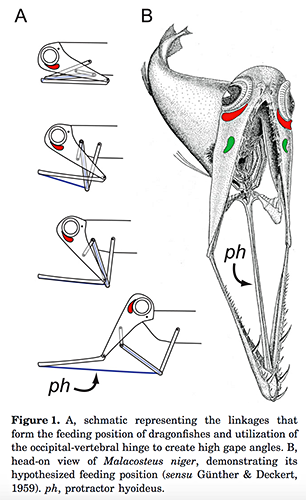
And there’s a lot to unpack here with the anatomy of this one.
First of all, there’s the fact that its entire head can hinge away from its body, gaping enormous jaws with long fang-like teeth.
The bottom of its lower jaw has no skin membrane connecting the two sides, attached to the rest of its bizarre head only by the hinges and a single exposed muscle, reducing water resistance so it can shoot its trap-jaws out extra fast to snare prey.
From Kenaley, C. P. (2012). Exploring feeding behaviour in deep-sea dragonfishes (Teleostei: Stomiidae): jaw biomechanics and functional significance of a loosejaw. Biological Journal of the Linnean Society, 106(1), 224-240. doi.org/10.1111/j.1095-8312.2012.01854.x
Once it catches something it retracts its head, and several sets of pharyngeal teeth further back grab hold of its prey and direct it down its throat.
(Let me remind you that this isn’t an early April Fools joke. This thing is completely real.)
In addition to all that anatomical weirdness, it’s also one of the only deep-sea fish that can both see and produce red-colored light. Most creatures living at that depth have lost the ability to see red since that frequency doesn’t penetrate so far down through water, but the stoplight loosejaw has evolved to take advantage of that by using bioluminescent red light as its own personal night vision goggles.
Using large red photophores under each eye, it can shine a spotlight out ahead of itself and see other deep-sea animals all clearly lit up, while remaining completely invisible to both them and any nearby larger predators. It’s able to perceive the color red thanks to a pigment in its eyes modified from chlorophyll, a visual setup unique to this fish and not known from any other vertebrate.
It also has a smaller green photophore further down on its head – inspiring its common name thanks to the resemblance to traffic lights – and many smaller blue and white ones over its head and body.
So, with its highly specialized jaws and ability to see things other deep-sea animals can’t, the stoplight loosejaw must be hunting something pretty impressive, right?
And as it turns out, it eats… plankton.
The vast majority of its diet appears to be copepods, small zooplanktoic crustaceans that are incredibly common in the waters the loosejaw inhabits. It may simply be “snacking” on such a convenient food source in-between rare encounters with larger prey – but it may also be getting the chlorophyll-based pigment needed for its night vision from eating them.
Much like Platybelodon from a few entries back, chalicotheres look like a fictional creature design rather than something that actually existed.
These animals were odd-toed ungulates related to modern horses, tapirs, and rhinos, who ranged across Africa, Eurasia, and North America for a large chunk of the Cenozoic. Instead of hooves they had large claws on their feet, and they appear to have occupied the same sort of ecological niche as ground sloths or therizinosaurs – sitting or rearing up on their hind legs to browse on high vegetation, using the hook-like claws on their forelimbs to pull down and strip branches.
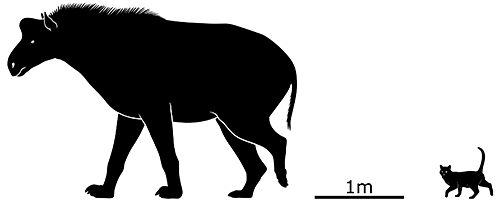
Tylocephalonyx skinneri here was one of the latter group, known from the Miocene of North America about 16-13 million years ago. Standing about 2m tall at the shoulder (6’6″), it had the same sort of chunky body as other schizotheriines and walked around with its large front claws held up to keep them raised away from the ground.
But there was also an unusual feature on its otherwise rather horse-like head – a large bony dome on top of its skull, like a mammalian version of a pachycephalosaur.
It probably used its dome in the same way as the dinosaurs it convergently resembled, headbutting or flankbutting in fights with each other.
With their odd-looking skulls, long tusks, and their noses and upper lips modified into tentacle-arm-like trunks, modern elephants are the sort of animal that would seem completely unbelievable if we only had fossils of them.
But not nearly as strange as some of their ancient relatives.
Platybelodon is probably the most famously weird member of the proboscideans (the group that contains both modern elephants and their extinct cousins), looking like some sort of deliberately outrageous speculative creature design.
Living during the mid Miocene, around 15-4 million years ago, several different species of Platybelodon ranged across Africa, Europe, Asia, and North America, with Platybelodon grangeri here known from abundant fossils in Asia.
These strange-looking proboscideans stood around 2.2m tall (7’3″) and had fairly standard elephant-like bodies, but also heads with bizarre-looking elongated lower jaws that ended in a wide flat shovel-like shape tipped by two flat tusks, leading to their nickname of “shovel-tuskers”.
It was originally interpreted as a swamp-dwelling animal using its weird jaw to scoop up soft aquatic vegetation, with a fairly short flat trunk. But more recent studies of the wear patterns on its teeth suggest it actually used them more like a scythe than a shovel, cutting through tough grasses and branches – a feeding style that would also require it to have a much more modern-elephant-like trunk, using it to hold on to plants while it was sawing through them.









{kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}