Fossils of cambroernids were first discovered in the early 1900s, but these Paleozoic animals were so confusing that for a long time their evolutionary relationships were a mystery.
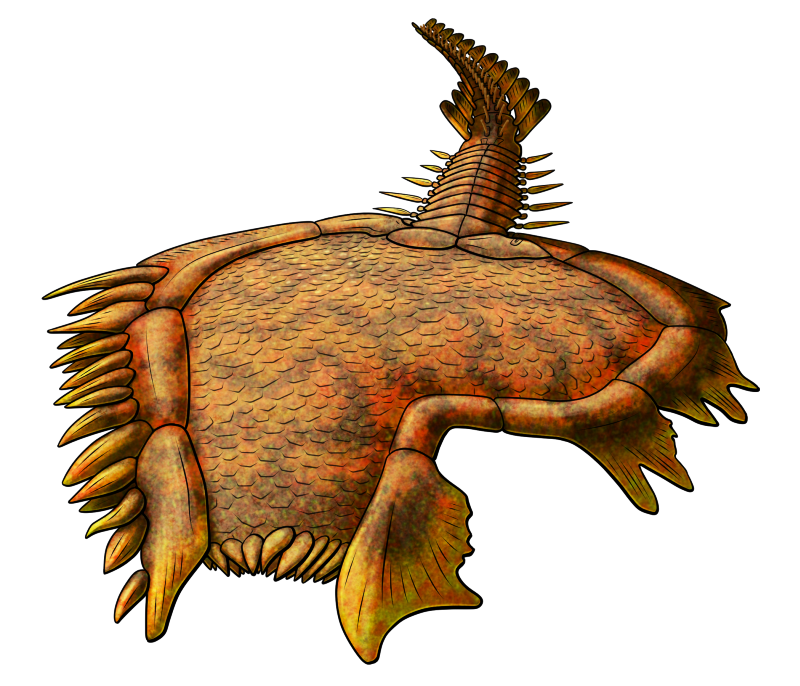
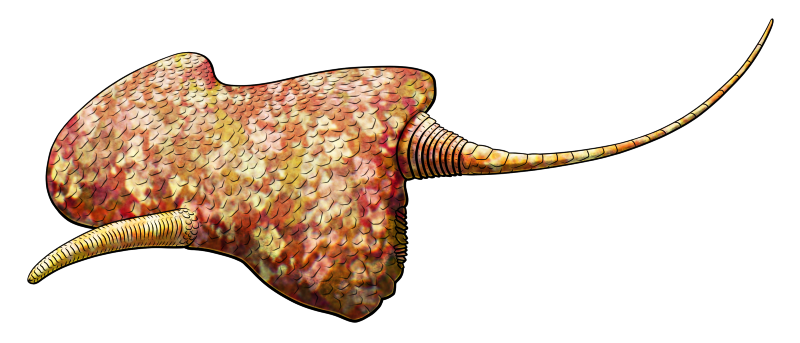
Discophyllum peltatum lived during the late Ordovician, about 458-448 million years ago, in what is now New York, USA. Up to around 11cm in diameter (~4.3″), it was one of the disc-shaped cambroernids – a lineage known as eldoniids – with a shallow domed body containing a clockwise-coiling sac and delicate feeding tentacles around its mouth.
Its disc would have been tough but flexible, containing numerous supporting radial structures that were probably part of a fluid-filled hydrostatic skeleton, giving it an almost-radially-symmetric body plan superficially resembling a jellyfish.
The lifestyle of eldoniids is still uncertain, but they seem to have mostly sat on the seafloor, possibly extending their tentacles out from under their discs to grab nearby food.
Early echinoderms seem to have gone through an asymmetrical phase before starting to evolving their characteristic radial symmetry. The first truly radial forms had three-way symmetry, but soon a group called the edrioasteroids upped that count to five.
First appearing in the fossil record around 525 million years ago in the early Cambrian, edrioasteroids were mostly shaped like discs or domes, and were immobile filter-feeders that lived permanently attached onto surfaces like the seafloor or the shells of other animals. Unlike most modern echinoderms their pentaradial symmetry was actually created by taking a tri-radial body plan and forking two of their arms near the bases to create a total of five.
Thresherodiscus ramosus
Thresherodiscus ramosus was an unusal edrioasteroid that lived in the shallow seas of what is now central Canada during the late Ordovician, around 460-450 million years ago. Up to about 4cm in diameter (~1.6″), its arms split additional times at irregular intervals, creating a complex asymmetrical branching pattern across its upper surface.
The tips of its arms protruded slightly over the rim of its body, and along with the erratic extra branching this may have been an adaptation to increase its food-gathering surface area.
Another group of early pentaradial echinoderms known as the blastozoans were characterized by erect feeding appendages called brachioles. But some blastozoans abandoned their five-way symmetry in favor of much stranger arrangements, sometimes having as few as two arms – and, in some cases, two mouths.
Amygdalocystites radiatus
Known from the same general area and time period as Thresherodiscus, Amygdalocystites radiatus was part of an Ordovician-to-early-Silurian lineage known as paracrinoids, which attached their irregularly-shaped bodies to the seafloor via a stem.
About 5cm long (~2″) Amygdalocystites had just two asymmetric arms forming “food grooves” along its upper edge, each lined with numerous long brachioles along just one of their sides. It probably orientated itself so its body was facing down-current, which would have created eddies that brought suspended food particles within easier reach of its brachioles.
Represented today by starfish, brittle stars, sea urchins, sea cucumbers, and crinoids, the echinoderms have a characteristic five-way radial symmetry that makes them barely even recognizable as bilaterians. Their true ancestry is only revealed by their genetics and their larvae, which still retain bilateral symmetry – and the way they metamorphose into adults is bizarre, essentially growing a whole new radial body from within the left side of their larval body.
(Sea cucumbers and sand dollars are superficially bilateral as adults, but evolved this secondarily on top of their existing radial symmetry. And some adult echinoderms like starfish also seem to retain a little bit of “behavioral bilaterism”, generally preferring to move with a specific arm always acting as their “front” end.)
The first known echinoderms appeared in the fossil record during the early Cambrian, about 525 million years ago, but the common ancestor of the whole group probably actually originated a few tens of millions of years earlier in the mid-to-late Ediacaran. Early echinoderms seem to have started off as flattened animals that sat on the seafloor filter-feeding, and with this largely immobile way of life their bodies started to shift into asymmetry, no longer constrained by the locomotory advantages of being bilaterally symmetric.
In fact, for these early sedentary filter-feeders being radial was actually much more advantageous, able to distribute sense organs all around their bodies and grab food from any direction without having to reposition themselves, converging on the lifestyle of non-bilaterian cnidarian polyps. The evolutionary transition from bilateral to asymmetrical to pentaradial seems to have happened incredibly quickly during the Cambrian Explosion, and all modern echinoderms probably evolved from a group called the edrioasteroids, maintaining their new base body plan even when they later began taking up more mobile lifestyles again.
But during the process of all that some very alien-looking lineages split off at various stages of anatomical weirdness.
Stylophorans had asymmetrical bodies with a single feeding arm at the front, and varied from irregular boot-like shapes to almost bilateral heart shapes depending on their specific ecologies. The highly asymmetrical forms were probably spreading their weight out over soft soupy mud in quiet waters, while the more bilateral forms may have been more streamlined to deal with stronger water currents.
Sokkaejaecystis serrata
Sokkaejaecystis serrata was a stylophoran that lived during the late Cambrian, about 501-488 million years ago, in what is now South Korea. It was tiny, only about 1cm long (~0.4″), and its boot-shaped body was surrounded by spines and flanges that spread out its surface area and probably also made it much more awkward for small predators to attempt to eat.
Meanwhile the solutes started off as immobile animals living attached to the seafloor via a stalk-like appendage. But fairly early in their evolution they switched to a more active mode of life, modifying their stems into tail-like “steles” that were used to push themselves along.
Maennilia estonica
Maennilia estonica lived in what is now Estonia during the late Ordovician, about 450 million years ago. It was quite large for a solute at about 12cm long (~4.7″), with a sort of vaguely-trapezoidal body, a short feeding arm, and a long thin stele.
Both of these strange early echinoderm lineages were surprisingly successful, surviving for a good chunk of the Paleozoic Era alongside their more familiar radial relatives. The solutes lasted until the early Devonian about 400 million years ago, and the stylophorans continued all the way into the late Carboniferous about 310 million years ago.
It was part of an extinct Paleozoic echinoderm lineage known as edrioasteroids, which lived attached to the seabed or on hard surfaces like the shells of other marine animals, using the tube feet on their five arms to catch food particles from the water around them.
Living during the Silurian, about 435 million years ago, in what is now Quebec, Canada, it stood around 3-4cm tall (1.2-1.6″), firmly anchored into the seafloor sediment by a bulbous sac-shaped base. Its long stalked body was somewhat flexible, and it was able to partially contract the top feeding region down under a “collar” of large scale-like armor plates.
Known from the Emu Bay Shale fossil deposits in Kangaroo Island, South Australia (~514 million years ago), Nesonektris was one of the larger known vetulicolians, growing to at least 17cm long (~6.5″). Like most of its relatives it had a large streamlined forebody with a mouth opening at the front, and no obvious appendages or sensory structures. A groove down each side may have housed gill openings, and a segmented flexible tail provided propulsion for swimming.
Very little is known about the ecology of these animals. They were clearly adapted for active swimming in the water column, and may have filter-fed on plankton – but some other vetulicolians have been found preserved with their guts full of seafloor sediment, suggesting some sort of detritivorous lifestyle instead.
Their evolutionary relationships are also still uncertain, but preservation of what appears to be a notochord in Nesonektris suggests that vetulicolians may have been part of the chordate lineage, possibly close relatives of tunicates.
Vetulicolians were a group of odd Cambrian animals known from between about 520 and 505 million years ago. The front half of their bodies were large and streamlined, with a prominent mouth, no eyes, and five pairs of openings that seem to have been gills, with some species having a rigid exoskeleton-like carapace. Their back half was slender, segmented, and flexible, and functioned as a tail for swimming, giving them an overall appearance like alien tadpoles.
Their evolutionary affinities have been problematic for a long time, but evidence of a notochord in some specimens suggest they were probably related to the chordates in some way. Sometimes they’re considered to represent their own phylum, but they might also be stem-chordates or stem-tunicates.
Vertebrates are by far the most numerous and diverse group of chordates today, with over 65,000 known species including fish, amphibians, reptiles, birds, and mammals. Genetic studies show that they’re closely related to the weird bag-like tunicates, and their shared common ancestor was probably something lancelet-like.
Cambroernids were a bizarre group with branching feeding tentacles and a gut enclosed in a coiled sac. They came in a range of forms from worm-like to cup-like to disc-shaped, and despite their fossils being known since the early 1900s their evolutionary affinities were a longstanding problem. Various species had been interpreted in the past as sea cucumbers, jellyfish, tunicates, gnathiferans, or lophophorates, but in recent years they’ve been recognized as all being related, and linked to the ambulacrarians.
And it’s still not entirely clear where in that group they actually belong. They were probably a weird early stem lineage, but they might also be early stem-hemichordates or stem-echinoderms.