
“Insectivora” was a wastebasket taxon so bad it had to be revised multiple times, but there’s another particularly infamous case in mammal taxonomy that’s still in the process of being resolved – the “condylarths“.
This group was first created in the early 1880s, during the Bone Wars, and initially was just a subgroup of odd-toed ungulates containing only the phenacodontids. But just a few years later Condylarthra was promoted up to its own order, and groups like the periptychids and hyopsodontids were added in too.
Then over the next few decades various groups were added and removed from the condylarths, most notably with the mesonychids and arctocyonids being brought in from their previous position with the creodonts.
By the mid-20th century the condylarths had become a big convenient dumping ground for any and all “primitive” ungulate-like mammals that didn’t easily fit into any modern groups, ranging in age from the early Paleocene through to the early Oligocene. But it soon became apparent that they had the same problem as the “insectivores” – there weren’t really any unique anatomical features that united all these animals together.
They generally had rounded-cusped molar teeth and hoof-like toes, but they also had rather generalized “primitive mammal” features and a diverse range of ecologies. Some were small herbivores, but others were coati-like or dog-like omnivores, and some were even bear-sized carnivores.

Bottom row: Ectoconus ditrigonus (periptychid), Mesonyx obtusidens (mesonychid)
It wasn’t even clear how the various different condylarth groups were actually related to each other. The best guess was that arctocyonids had arisen from within the “insectivores”, with a Protungulatum-like form as the common ancestor of all the other condylarths. Where exactly modern ungulates had then evolved from within the condylarths was also still uncertain.
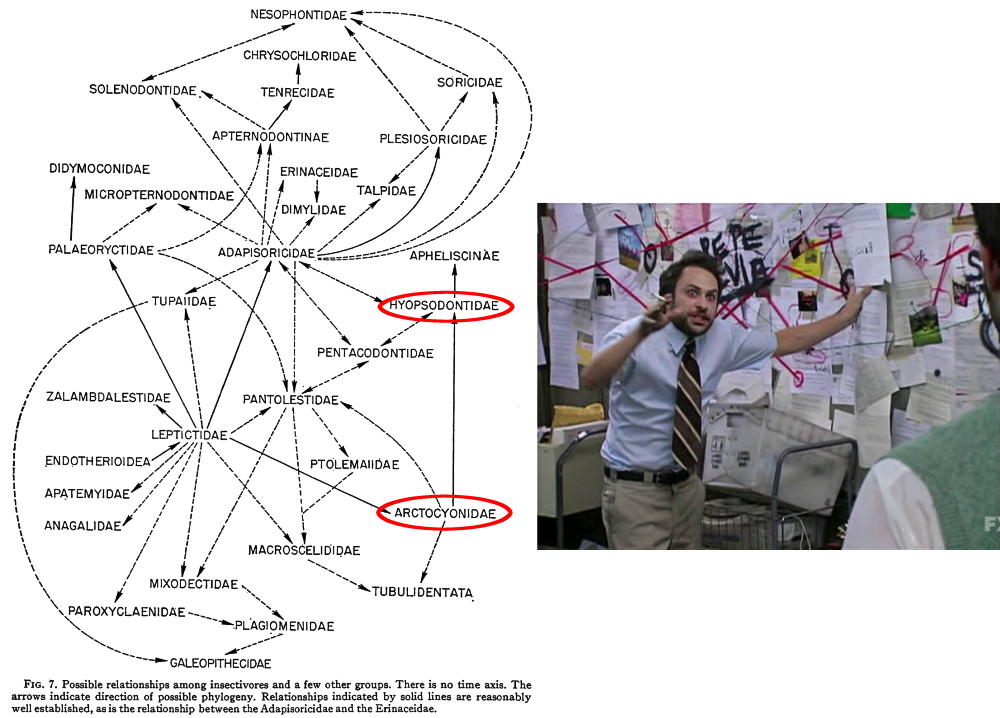
(Image sources: http://hdl.handle.net/2246/358 & https://knowyourmeme.com/memes/pepe-silvia)
Cladistic analysis in the 1980s began to tackle the confusing pile of assorted condylarths, and showed that they weren’t the single ancestral source of all modern ungulates, but instead a loose collection of several unrelated groups from all over the ungulate evolutionary tree. Arctocyonids, periptychids, and hyopsodontids were placed as early “primitive” lineages, phenacodontids were loosely linked with the ancestors of odd-toed ungulates once again, and mesonychids were considered to be the ancestors of whales.
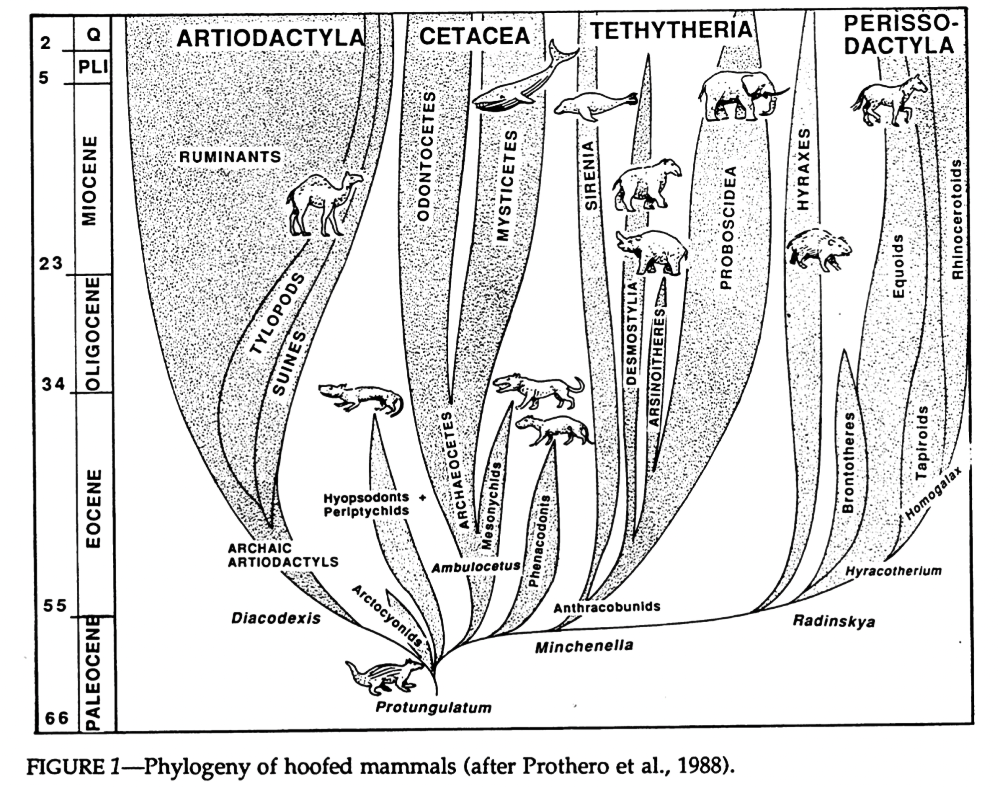
(Image source: https://doi.org/10.1017/S2475263000001343)
And, once again paralleling the mess of the “insectivores”, it wasn’t until genetic methods became available in the late 1990s that larger-scale ungulate relationships began to be properly resolved. The paenungulates (elephants, hyraxes, and sirenians), which had been traditionally considered to be a major branch of ungulates, were removed entirely and reclassified as afrotheres. And, along with some new fossil discoveries, whales were recognized as having actually evolved from within the even-toed ungulates instead of from mesonychids.
This shake-up threw the still-problematic “condylarth” classifications back into question – with some “condylarths” turning out to also be afrotheres instead of true ungulates.
Today the actual relationships of the main “condylarth” ungulate families are still in the process of being figured out, and there’s a lot of remaining uncertainty and disagreement about them.
Phenacodontids seem to have mostly maintained their traditional position as early odd-toed ungulates, and hyopsodontids may potentially be part of this group too – possibly as members of the hippomorph lineage, closely related to horses and brontotheres. Arctocyonids might be a wastebasket themselves, with some studies finding them to be a mix of several different archaic ungulate lineages. Periptychids may have links to the even-toed ungulates. The mesonychids, meanwhile, are now generally considered to be a separate order from the traditional “condylarths”, and may be either an early branch of the even-toed ungulates or much more basal ungulates closely related to the “arctocyonids”.
Since the term “condylarth” no longer has any real taxonomic meaning some paleontologists have proposed replacing it with “archaic ungulate” to distance from the historical messiness of the old name. But this hasn’t really caught on, and many papers still use “condylarth” in a very loose sense to refer to an “evolutionary grade” of early ungulates of unclear evolutionary affinities.
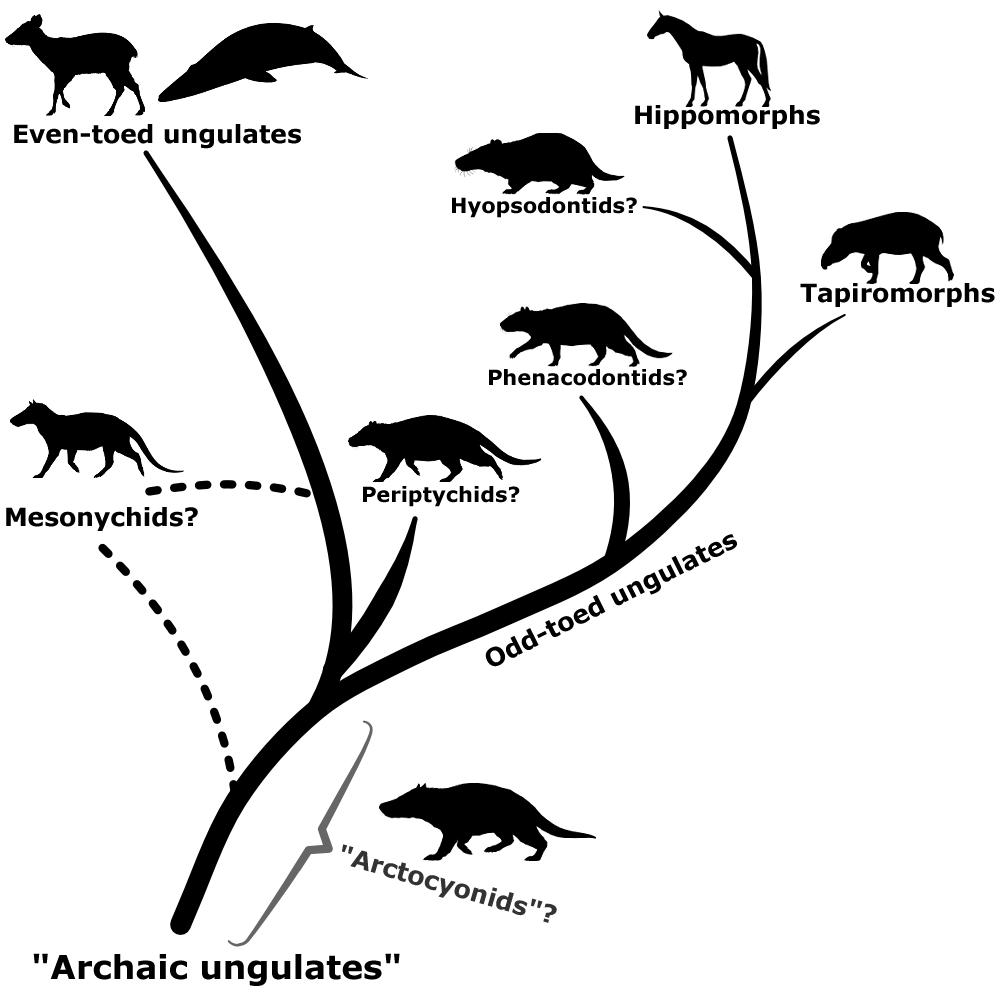
And while that’s the last main entry for this month, we’re not quite done yet. There’s still one weekday left in October, and after digging through so many taxonomic garbage cans there’s only one place we can go now.
…See you in the trash heap.








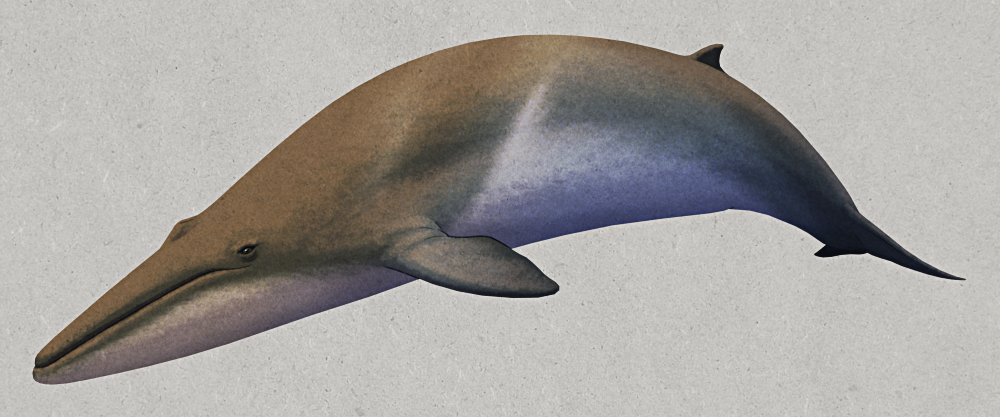

{kind=link}