

Manipulonyx reshetovi was an alvarezsaurid theropod dinosaur that lived during the Late Cretaceous, about 70 million years ago, in what is now Mongolia.
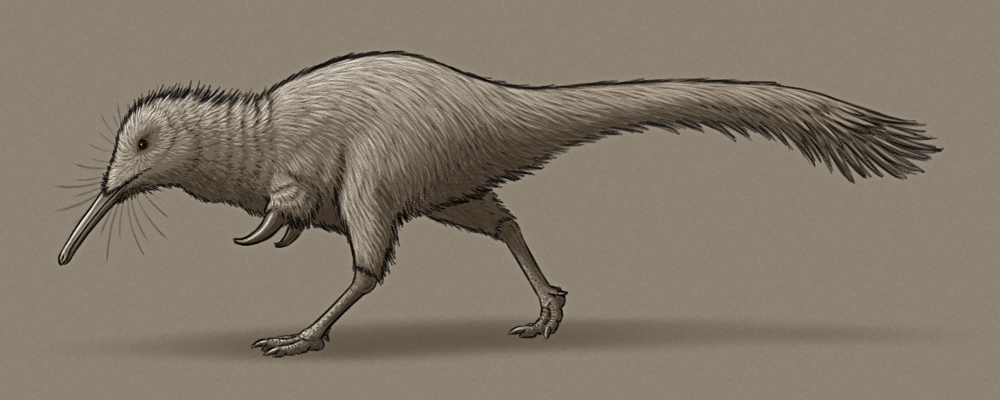
Although only known from a partial skeleton, based on its close relatives it would have been about 50cm long (~1’8″), with a narrow snout, a coat of fluffy down-like feathers, long slender legs, and short stout arms with massively enlarged thumb claws.
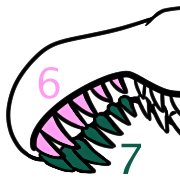
Notably the fossil specimen has the best preserved hands known from the parvicursorine subfamily of alvarezsaurs, revealing a surprising arrangement of three ossified conical spikes around the thumb. These hand spikes were positioned pointing forwards, backwards, and inwards, and may have improved the ability to grip with just one functional finger.
“Spike-like” hand bones were also previously reported in 2009 in another parvicursorine, Ceratonykus, suggesting that these structures weren’t unique to Manipulonyx.
The paper describing Manipulonyx proposes that alvarezsaurs were ovivores, using their hands to carry and then break open eggs, but I’m personally very skeptical of this idea. I think the myrmecophagy interpretation for these little dinosaurs is still much more likely – they probably did opportunistically raid eggs when they could get them, like many modern predators do, but being specialized for just egg-eating is a big claim that definitely needs more evidence.
Continue reading “Manipulonyx”


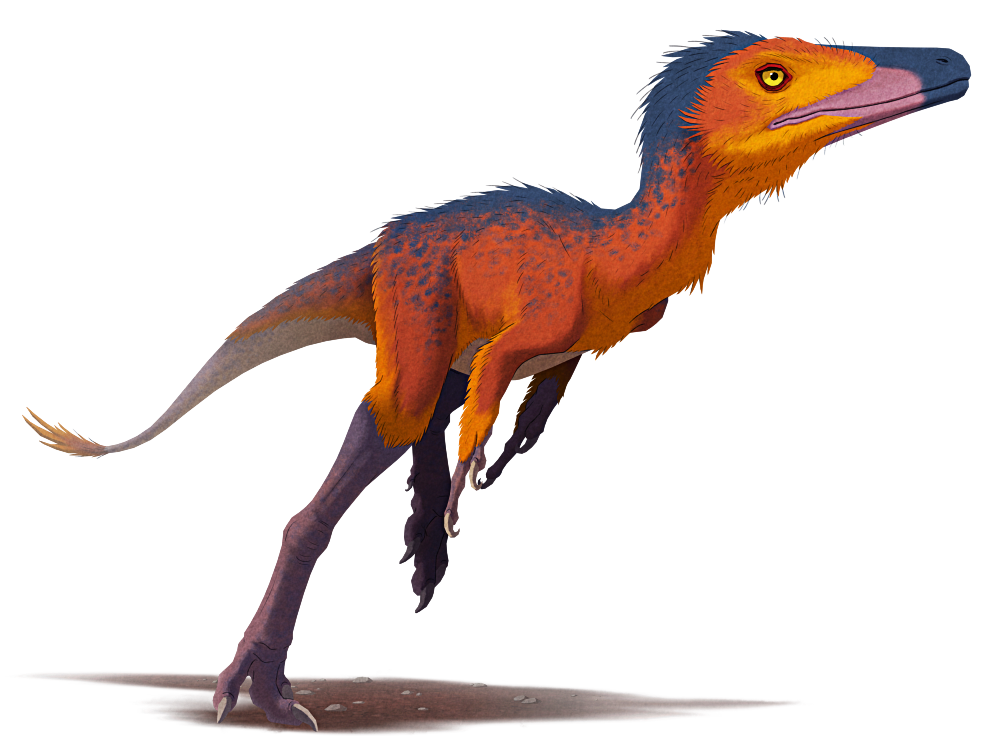







{kind=link}