

At the end of the Miocene, about 6 million years ago, the movement of tectonic plates caused the narrow connection between the Mediterranean Sea and the Atlantic Ocean to close — and over the next several hundred thousand years the Mediterranean dried up almost completely.
5.3 million years ago the Strait of Gibraltar formed, re-establishing the connection with the Atlantic, and the Mediterranean refilled incredibly rapidly, possibly in as little as two years. As a result, various species that had colonized across the dried-out Mediterranean from the continental mainland were left stranded out on islands that had been re-formed throughout the sea.
And on what is now Menorca a population of rabbits found themselves isolated, with little competition and no large terrestrial predators.

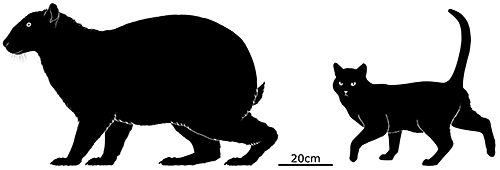
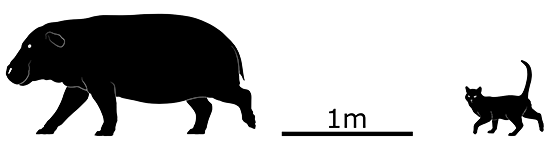
Nuralagus rex was the evolutionary result, an enormous rabbit 50cm tall at the shoulder (1’8″). It was heavily built with a stiff spine — making it unable to hop — and had weaker senses than its ancestors, with small eyes and stubby ears. It would have been a slow-moving animal ambling around the scrublands of Menorca, digging for its main foods of roots and tubers.
It’s unclear what happened to this big bunny, but it seems to have disappeared around the end of the Pliocene, about 3-2.5 million years ago. Possibly the onset of global cooling at the beginning of the Pleistocene ice ages changed the climate too quickly for it to adapt to, or the dropping sea levels that connected Menorca with nearby Mallorca introduced new competition from the other island that Nuralagus couldn’t cope with.

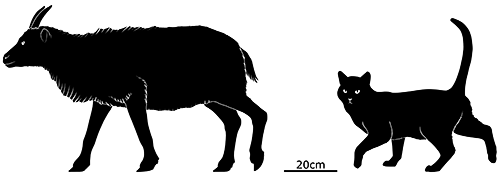
And one of the animals that spread into Menorca from Mallorca was Myotragus.
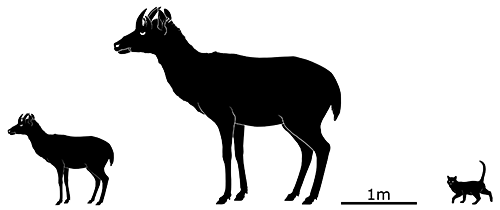
About the same size as the giant rabbits, at 50cm tall (1’8″), Myotragus balearicus was a close relative of modern sheep that had undergone dwarfing on Mallorca since its ancestors’ isolation 5.3 million years ago.
It was possibly one of the most unusual mammals ever, with a combination of features not seen anywhere else. Its snout was relatively short and rabbit-like, with ever-growing front teeth in its lower jaw, and its eyes faced directly forward, giving it stereoscopic vision more like a primate than a herbivore. Its brain and sense organs were highly reduced compared to its ancestors, its legs were shorter and stockier, and its feet had lost much of their flexibility, making it unable to run or jump.
But strangest of all was its metabolism, as indicated by growth lines in its bones. It was essentially cold-blooded, functioning more like a reptile than a mammal, growing at a slow rate that varied or even stopped entirely depending on the conditions of its environment. It would have taken around 12 years for it to reach maturity, an incredibly long time for its size, and so it would have reproduced very very slowly — but this metabolic strategy also allowed it to conserve a lot of energy and survive long periods of scarce food availability.
Myotragus‘ extreme weirdness obviously worked to its advantage, because it was quite common on Mallorca-Menorca and unlike many of the other native species it survived through the much colder drier conditions of the Pleistocene.
It was still around in the Holocene when humans reached the islands in about 3000 BCE, and this slow-moving sluggish goat-like animal with a rather uncanny face must have been a very strange sight to them.
There was a hypothesis that the early settlers actually attempted to domesticate Myotragus, based on remains found in caves with what appeared to be trimmed horns. But in recent years this has been disputed, since some of the “trimmed” horns pre-date human arrival and may be better explained as the result of Myotragus individuals chewing on the bones of others for the mineral content.
Shortly after the arrival of humans Myotragus and the other remaining endemic mammals on the islands (a giant dormouse and a giant shrew) disappeared entirely, suggesting that a combination of hunting, competition from introduced livestock, and predation from dogs was finally too much for these strange sheep to handle.













{kind=link}
{kind=link}
{kind=link}