

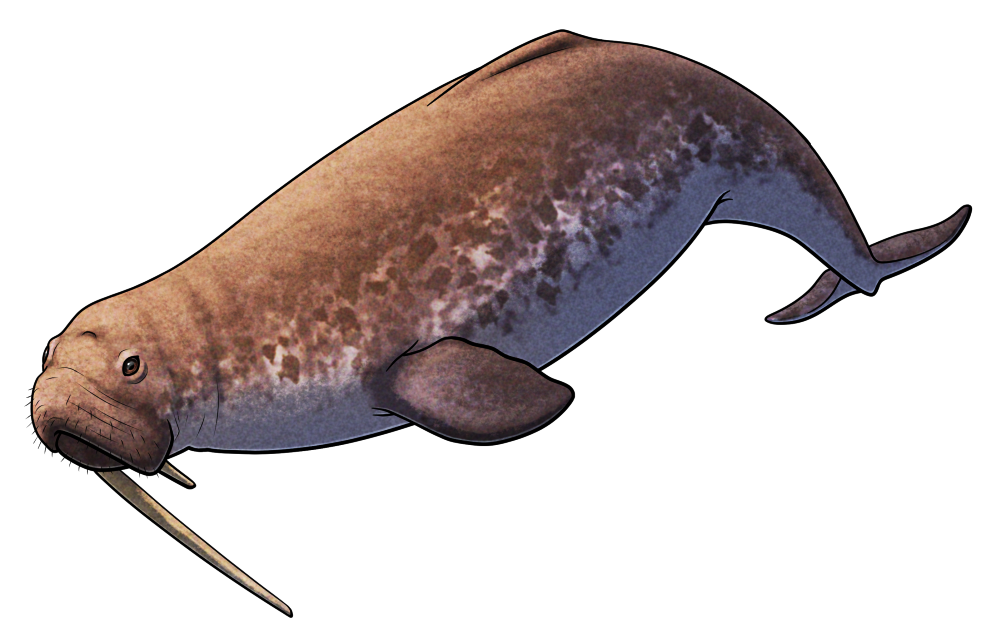
Aureia rerehua was a toothed whale that lived during the late Oligocene and early Miocene, around 23-22 million years ago, in shallow coastal waters covering most of what is now Aotearoa New Zealand.
It was closely related to the waipatiids – a group traditionally classified as platanistoids (the South Asian river dolphin lineage), but more recently proposed as instead representing a separate earlier branch of the toothed whale evolutionary tree.
About 2m long (~6’6″), Aureia had distinctive tusk-like teeth that splayed outwards from its snout, interlocking when its mouth was closed. Along with a flexible neck and its fairly delicately-built skull and jaws, this suggests it was specialized for catching small prey in a “fish trap” of teeth, a unique feeding strategy for a toothed whale.
Along with different feeding specializations in other close relatives like Nihohae, Aureia shows us how multiple species of these ancient Aotearoan cetaceans were able to coexist in the same place and time by diversifying into different novel ecological niches.
Continue reading “Aureia”









{kind=link}
{kind=link}
{kind=link}