Brachiopods (also known as “lamp shells”) superficially look very much like bivalves, but these two groups aren’t very closely related to each other – although they’re both lophotrochozoans, their last common ancestor probably lived sometime in the Ediacaran at least 560 million years ago, and their similarities in appearance are due to convergent evolution.
The two valves of their shells are also arranged differently. Bivalve shells grow on their left and right sides and are usually symmetrical, but brachiopods form their shells from the upper and lower surfaces of their bodies.
As a result brachiopod shells are usually unequal in size and shape but have their own plane of bilateral symmetry down the center – but some of them still managed to become asymmetrical anyway.
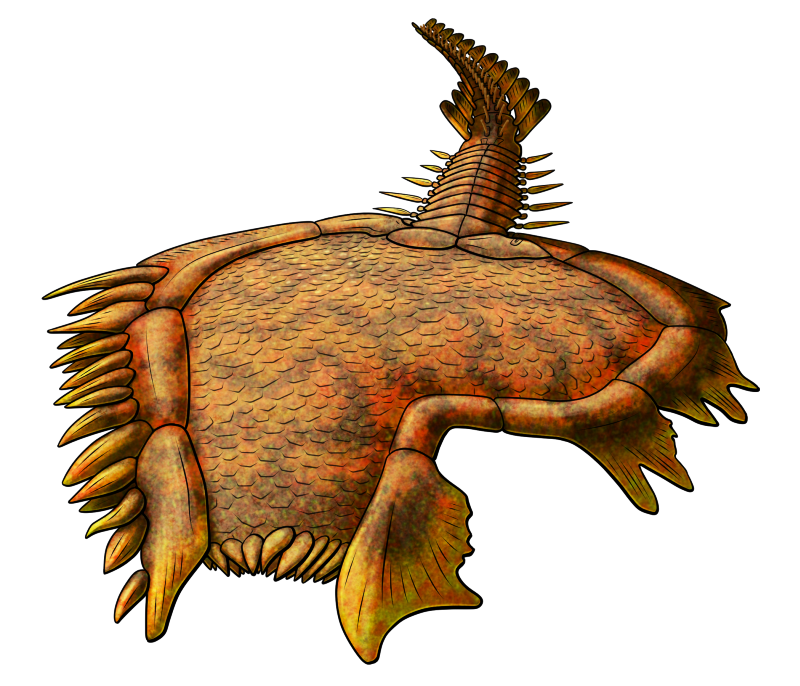
Torquirhynchia inconstans lived during the Late Jurassic, about 161-145 million years ago, in the warm shallow seas that covered what is now Europe and Iran. Around 3cm across (~1.2″) it had a strongly ridged shell with an asymmetrical closing edge, positioned high on one side and low on the other.
This unusual uneven arrangment is thought to be an adaptation to living on soft sediments. Asymmetrical brachiopods like Torquirhynchia may have lived with one side of their body mostly buried into the seafloor, and twisted their shell edges so the still-exposed half was raised up to better function for water circulation and filter-feeding.
But some bivalves break that arrangement, developing asymmetrical valves that can be massively different in size and shape.
Gryphaea arcuata was an oyster that lived during the Early Jurassic, about 200-174 million years ago, in the warm shallow seas that covered what is now Europe and eastern Greenland. Around 6cm long (~2.4″), its left valve was thick and strongly convex and curled, while the right valve was relatively thin and slightly concave forming a “lid”.
The gnarled curled claw-like shape of Gryphaea fossils led to them being colloquially known as “devil’s toenails” in some of the regions where they’re commonly found, with folk beliefs that they had the power to prevent joint pain.
Their shape was actually an adaptation to living on very soft seafloor sediments. The larger curled valve acted sort of like a boat on the soupy mud, supporting the Gryphaea‘s weight and preventing it from sinking.
With its bizarre boomerang-shaped skull, Diplocaulus is probably the most iconic ancient amphibian. (It even inspired the design of the pokémon Dragapult!) It was part of the lepospondyls, a diverse collection of early tetrapods mainly found in Europe and North America between the Early Carboniferous and the mid-Permian, about 350-270 million years ago.
But one species hung on a bit longer into the late Permian, about 259-254 million years ago, and this late-surviving lepospondyl was perhaps the oddest of them all.
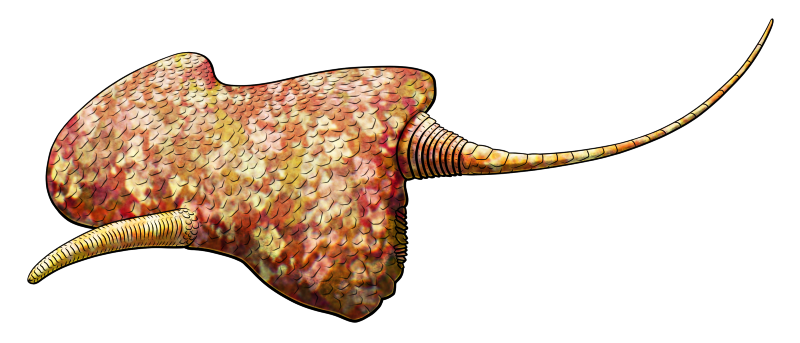
Diplocaulus minimus was the only lepospondyl known from the supercontinent of Gondwana, found in what is now Morocco in northern Africa. About 70cm long, around half of which was its long tail, it had the distinctive elongated skull of a diplocaulid – but in a bizarrely asymmetrical shape.
The left prong of its skull was long and tapering, but the right was shorter and more rounded. This doesn’t seem to have been due to individual deformity or distortion of the fossil material, since more than one skull has been found with the same features, but the reason for such a striking amount of asymmetry in this species is unknown.
Diplocaulids’ head shapes are thought to have acted as hydrofoils, providing lift while they were swimming. Perhaps Diplocaulus minimus‘ much more wonky skull means this species wasn’t relying on that hydrodynamic function as much as its relatives, and something else was going on with its ecology.
The spiral-coiled shells of snails are their most familiar feature, giving them obvious external asymmetry, but gastropods are also defined by a specific type of internal asymmetry known as torsion.
Torsion is an anatomical process that occurs during larval development, and involves rotating their internal organs, mantle, and shell a full 180° relative to their head and muscular foot. This twists their gut into a U-shape, knots up their nervous system, and brings their respiratory organs and anus up close to their head.
And we still don’t really know why they do it.
One idea (the “rotation hypothesis”) is that it originated as a defensive function after early gastropods began developing their spiral shells. The shell opening may have originally been positioned at early gastropods’ rears, meaning they retracted their bodies back-end-first leaving their heads and sensory structures still vulnerable – but twisting the shell around would allow them to pull their front end in faster instead.
A competing idea (the “asymmetry hypothesis“) instead proposes that the shape of the coiled shell restricted the gills of early gastropods, which may have originally been positioned in mantle cavities on each side of their bodies. In response to this they developed a single larger gill cavity on just one side of their body, and then gradually expanded and rotated this asymmetric feature around to the front for better aeration.
In either case this resulted in some of the rest of their anatomy “coming along for the ride”. And regardless of whatever the original evolutionary advantage of torsion actually was, it made gastropods incredibly successful – they’re a massively diverse group, second only to the insects in terms of sheer number of species, and today they’re found all over the world in almost every habitat from deep sea trenches to high mountain elevations.
Spinyplatyceras arkonense
Spinyplatyceras arkonense lived in what is now Ontario during the mid-Devonian, about 391-385 million years ago. Around 5cm long (2″), it was part of a group of Paleozoic marine snails known as platyceratids, which were probably related to either modern limpets or neritomorphs.
Platyceratids seem to have had a unique parasitic relationship with crinoids, attaching themselves to the top of the host’s body and using their radula to drill into them, either robbing food directly from the crinoid’s gut or feeding on its other internal organs.
The long spines on Spinyplatyceras‘ shell probably helped to deter predators. In an interesting case of coevolution the crinoid hosts of some platyceratids developed their own defensive spines, too – and it seems this wasn’t to prevent the snails from infesting them, but to also discourage the snails’ predators. These crinoids may have been frequently indirectly injured during snail-eating predators’ attacks, and it might have actually “cost” them less to keep enduring an infestation than to deal with the collateral damage of the snails being removed.
The genus Walliserops was one of the weirdest-looking trilobites, covered in numerous pointy spines and sporting a large three-pronged “trident” on the front of its face.
They also had some degree of asymmetry in their bodies. Their tridents often didn’t fork evenly, and their long forehead spines curved off to one side – possibly so they could lift their heads up without stabbing themselves in the back.
Walliserops hammii lived in what is now Morocco during the early-to-mid Devonian, about 403-392 million years ago. Around 5cm long (~2″) It was one of the “short trident” species of Walliserops, and its chunky forehead spine curved particularly strongly to the right.
The function of these trilobites’ elaborate tridents is still poorly understood. But an unusual individual of the long-tridented species Walliserops trifurcatushas been found with a lopsided four-pronged trident, and since it was able to grow to full maturity the shape of the structure probably wasn’t absolutely vital for survival, suggesting it wasn’t used for feeding or sensory purposes.
The tridents may instead have been used for combat with each other similar to the horns of some modern beetles. However, these sorts of features are usually only seen in males, and there’s currently no definite evidence for any significant sexual dimorphism in trilobites.
(Although perhaps like ceratopsid dinosaurs their ornaments were just present in both males and females, being also useful for species recognition, visual display, and defense against predators.)
Fossils of cambroernids were first discovered in the early 1900s, but these Paleozoic animals were so confusing that for a long time their evolutionary relationships were a mystery.
Discophyllum peltatum lived during the late Ordovician, about 458-448 million years ago, in what is now New York, USA. Up to around 11cm in diameter (~4.3″), it was one of the disc-shaped cambroernids – a lineage known as eldoniids – with a shallow domed body containing a clockwise-coiling sac and delicate feeding tentacles around its mouth.
Its disc would have been tough but flexible, containing numerous supporting radial structures that were probably part of a fluid-filled hydrostatic skeleton, giving it an almost-radially-symmetric body plan superficially resembling a jellyfish.
The lifestyle of eldoniids is still uncertain, but they seem to have mostly sat on the seafloor, possibly extending their tentacles out from under their discs to grab nearby food.
Early echinoderms seem to have gone through an asymmetrical phase before starting to evolving their characteristic radial symmetry. The first truly radial forms had three-way symmetry, but soon a group called the edrioasteroids upped that count to five.
First appearing in the fossil record around 525 million years ago in the early Cambrian, edrioasteroids were mostly shaped like discs or domes, and were immobile filter-feeders that lived permanently attached onto surfaces like the seafloor or the shells of other animals. Unlike most modern echinoderms their pentaradial symmetry was actually created by taking a tri-radial body plan and forking two of their arms near the bases to create a total of five.
Thresherodiscus ramosus
Thresherodiscus ramosus was an unusal edrioasteroid that lived in the shallow seas of what is now central Canada during the late Ordovician, around 460-450 million years ago. Up to about 4cm in diameter (~1.6″), its arms split additional times at irregular intervals, creating a complex asymmetrical branching pattern across its upper surface.
The tips of its arms protruded slightly over the rim of its body, and along with the erratic extra branching this may have been an adaptation to increase its food-gathering surface area.
Another group of early pentaradial echinoderms known as the blastozoans were characterized by erect feeding appendages called brachioles. But some blastozoans abandoned their five-way symmetry in favor of much stranger arrangements, sometimes having as few as two arms – and, in some cases, two mouths.
Amygdalocystites radiatus
Known from the same general area and time period as Thresherodiscus, Amygdalocystites radiatus was part of an Ordovician-to-early-Silurian lineage known as paracrinoids, which attached their irregularly-shaped bodies to the seafloor via a stem.
About 5cm long (~2″) Amygdalocystites had just two asymmetric arms forming “food grooves” along its upper edge, each lined with numerous long brachioles along just one of their sides. It probably orientated itself so its body was facing down-current, which would have created eddies that brought suspended food particles within easier reach of its brachioles.
Represented today by starfish, brittle stars, sea urchins, sea cucumbers, and crinoids, the echinoderms have a characteristic five-way radial symmetry that makes them barely even recognizable as bilaterians. Their true ancestry is only revealed by their genetics and their larvae, which still retain bilateral symmetry – and the way they metamorphose into adults is bizarre, essentially growing a whole new radial body from within the left side of their larval body.
(Sea cucumbers and sand dollars are superficially bilateral as adults, but evolved this secondarily on top of their existing radial symmetry. And some adult echinoderms like starfish also seem to retain a little bit of “behavioral bilaterism”, generally preferring to move with a specific arm always acting as their “front” end.)
The first known echinoderms appeared in the fossil record during the early Cambrian, about 525 million years ago, but the common ancestor of the whole group probably actually originated a few tens of millions of years earlier in the mid-to-late Ediacaran. Early echinoderms seem to have started off as flattened animals that sat on the seafloor filter-feeding, and with this largely immobile way of life their bodies started to shift into asymmetry, no longer constrained by the locomotory advantages of being bilaterally symmetric.
In fact, for these early sedentary filter-feeders being radial was actually much more advantageous, able to distribute sense organs all around their bodies and grab food from any direction without having to reposition themselves, converging on the lifestyle of non-bilaterian cnidarian polyps. The evolutionary transition from bilateral to asymmetrical to pentaradial seems to have happened incredibly quickly during the Cambrian Explosion, and all modern echinoderms probably evolved from a group called the edrioasteroids, maintaining their new base body plan even when they later began taking up more mobile lifestyles again.
But during the process of all that some very alien-looking lineages split off at various stages of anatomical weirdness.
Stylophorans had asymmetrical bodies with a single feeding arm at the front, and varied from irregular boot-like shapes to almost bilateral heart shapes depending on their specific ecologies. The highly asymmetrical forms were probably spreading their weight out over soft soupy mud in quiet waters, while the more bilateral forms may have been more streamlined to deal with stronger water currents.
Sokkaejaecystis serrata
Sokkaejaecystis serrata was a stylophoran that lived during the late Cambrian, about 501-488 million years ago, in what is now South Korea. It was tiny, only about 1cm long (~0.4″), and its boot-shaped body was surrounded by spines and flanges that spread out its surface area and probably also made it much more awkward for small predators to attempt to eat.
Meanwhile the solutes started off as immobile animals living attached to the seafloor via a stalk-like appendage. But fairly early in their evolution they switched to a more active mode of life, modifying their stems into tail-like “steles” that were used to push themselves along.
Maennilia estonica
Maennilia estonica lived in what is now Estonia during the late Ordovician, about 450 million years ago. It was quite large for a solute at about 12cm long (~4.7″), with a sort of vaguely-trapezoidal body, a short feeding arm, and a long thin stele.
Both of these strange early echinoderm lineages were surprisingly successful, surviving for a good chunk of the Paleozoic Era alongside their more familiar radial relatives. The solutes lasted until the early Devonian about 400 million years ago, and the stylophorans continued all the way into the late Carboniferous about 310 million years ago.
It was a small worm-like animal about 1.6cm long (~0.6″), with bundles of long bristles along its sides and flap-like structures on its underside. Its back was also covered with sclerite armor arranged in a strangely asymmetrical fashion, with larger overlapping plates in the middle and numerous smaller cap-like sclerites distributed unevenly along each side.
Its discovery actually confirms an old prediction that lophophorates probably originated from armored worm-like animals, representing an evolutionary link between earlier free-living annelid-like forms and later immobile filter-feeding tommotiids.
It’s not known why the armor on Wufengella‘s back was so unevenly organized – but some of the later tube-like tommotiids also had weird symmetry going on, with forms like Eccentrotheca having irregular sclerites arranged in a spiral around their bodies.
Most animals are bilaterally symmetric, having body plans with mirrored left and right sides – which also allows them to have a defined head end, rear end, top side, and underside.
It’s not entirely clear what evolutionary advantage this type of symmetry gave to the first bilaterians, which would have been been small “simple” worm-like animals living sometime during the Ediacaran Period between 600 and 560 million years ago. The current generally accepted explanation is that it probably allowed for better active locomotion – clustering sense organs at the head end and directing body movement more efficiently towards food sources and away from threats.
…But not every bilaterian has stayed roughly symmetrical.
Over the last half-billion years or so some bilaterians have abandoned their roughly-mirror-image body plans in favor of something distinctly wonkier. Asymmetry has evolved multiple times in various different lineages, and so every weekday this month we’ll be looking at some examples.
And we might as well start way back near the beginning:
Strange Symmetries #01: Almost Bilateral
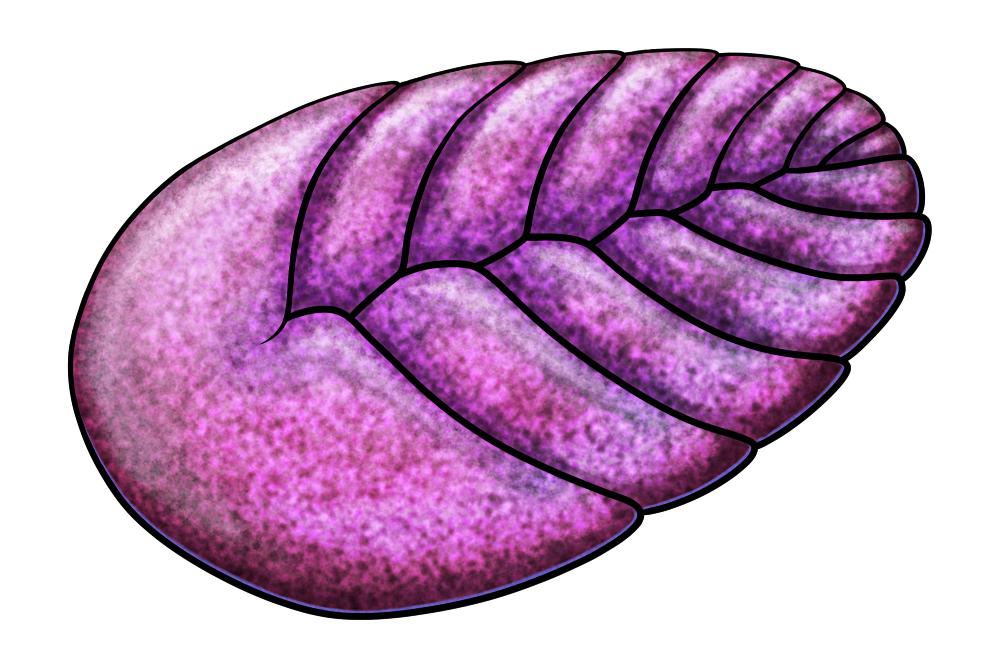
Living in the Ediacaran between about 567 and 550 million years ago, the proarticulatans were flattened rounded organisms with two rows of soft “quilted” rib-like segments (known as isomers) and sometimes a larger fused “head” section at the front. The left and right isomers weren’t perfectly mirrored, instead being offset from each other in a glide reflection pattern – but the presence of a clear central body axis suggests these animals may have had some sort of relation to the earliest bilaterians, possibly even being a very early stem group that was experimenting with a not-quite-totally-bilateral body plan.
Vendia sokolovi
Discovered in what is now northwest Russia, and dating to around 555 million years ago, Vendia sokolovi was a small proarticulatan measuring about 1.1cm long (0.4″). It had a rather small number of isomers compared to some of its relatives, only 7 per side, and seems to have had a simple digestive tract that branched into each isomer.
(The superficial resemblance to trilobites was coincidental – while we might not be entirely sure what these things were, we do at least know they weren’t closely related to early arthropods.)
Very little overall is known about these animals’ lifestyles. Trace fossils suggest they were able to move around, feeding on microbial mats on the seafloor, and they may also have been able to firmly stick themselves onto the spots they were currently grazing.











.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}