

Transcript for the text on the image under the cut:
Continue reading “Spectember #12: Cave Sheep”Spectember #11: Neotenic Echinoderms
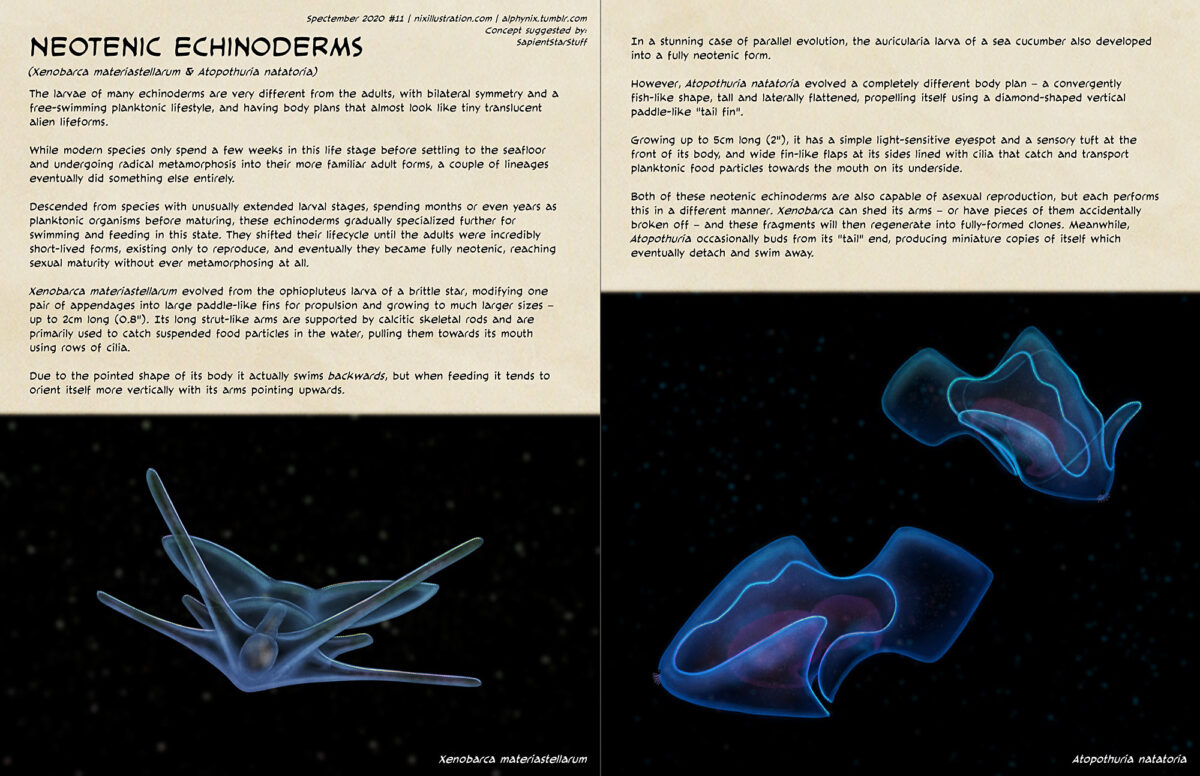
Transcript for the text on the image under the cut:
Continue reading “Spectember #11: Neotenic Echinoderms”Spectember #10: Marine Temnospondyls

Transcript for the text on the image under the cut:
Continue reading “Spectember #10: Marine Temnospondyls”Spectember Interlude: The Worlds of Wayne Barlowe
The After Man trilogy and The Future Is Wild might be the most well-known Earth-based works of speculative evolution, but for extraterrestrial life one of the most famous names is the American artist Wayne Douglas Barlowe.
Starting off illustrating the covers of science fiction novels, his first major project was Barlowe’s Guide to Extraterrestrials in 1979. Depicting alien species from various works of sci-fi in the format of a biological field guide, attempting to make them all look like real believable lifeforms, this book was a huge success at the time and eventually had a fantasy-themed sequel in 1996, Barlowe’s Guide to Fantasy.
But it was his 1990 book Expedition that helped define the alien side of the modern speculative evolution movement. Presented as a collection of field notes, journal entries, and paintings in his distinctive style, it chronicles a human crew’s visit to the planet Darwin IV in the year 2358, encountering a wide variety of strange organisms and trying to avoid having any lasting impact on the natural environment of this sometimes deadly planet. Some parts of the Darwinian species’ anatomy resembles Earth tetrapods, taking particular inspiration from the 20th century “alien prehistoric world” trope, but other elements are very different – all the animals appear to lack eyes, mostly sensing their surroundings using sonar and infrared receptor pits, there are no true jaws but instead various hollow proboscis “tongues”, there are flying forms using biological jet propulsion, and some species reach absolutely gigantic sizes compared to Earth life.
Expedition was adapted into a television documentary titled Alien Planet in 2005. Originally conceived as a nature film with little to no narration, the final product was more of a dramatization interspersed with interviews with various scientists and sci-fi filmmakers. Instead of a human presence on the planet, here the biological surveying is performed by autonomous drone-like probes with basic artificial intelligence.
Barlowe has also been involved in concept art for films, television shows, and videogames, with spec-evo relevant projects including the early design work for the creatures of Pandora in James Cameron’s Avatar, and the Barsoomian inhabitants of Mars in John Carter.
After being long out of print, a new edition of Expedition is due out this year (2020) from Echo Point Books: https://www.echopointbooks.com/art/expedition
Next week: more speculative dinosaurs!
Spectember Interlude: Mr. Dixon’s Wild Ride
The rhinogrades were the start of larger-scale speculative evolution works, but what truly popularized the genre to the general public are undoubtedly the works of Scottish geologist Dougal Dixon.
His 1981 book After Man was a “zoology of the future” using over a hundred speculative species to demonstrate the principles of evolution. Set 50 million years from now in the world of the Posthomic period, humans are long gone and the hardy survivors of a prior mass extinction have diversified and convergently evolved to fill vacant ecological niches, including rabbits becoming ungulate-like animals, penguins becoming giant whale-like creatures, and rats taking on the role of canid-like predators.
Some of the more outlandish creations have become infamous over the years and may seem cliché by modern standards – like the giant flightless predator bats – but when it was first published it was revolutionary, and there’s still a certain charm to it all even if the artwork isn’t quite as good as it could have been. Despite Dixon himself being a talented artist, the publisher chose to have all the final images created by other illustrators based on the original concepts. (And the updated 2018 edition goes as far as replacing a couple of images that the author was especially unhappy with.)
The success of After Man led to two “sequels” a few years later. In 1988 The New Dinosaurs took the premise of the K-Pg mass extinction never happening and used “an alternative evolution” to explain the geographical distribution of species. Inspired by the dinosaur renaissance, some of its fictional creations arguably predicted later paleontological discoveries – many species are depicted as extensively fuzzy (including ornithischians!) and aren’t too terribly shrinkwrapped for the time, and there are then-unknown concepts like small arboreal climbing theropods, giraffe-like ground pterosaurs, and semi-aquatic beaver-like mammals coexisting alongside the dinosaurs.
But at the same time much of it just hasn’t aged very well, with a lot of the paleontological basis of the The New Dinosaurs having become horrendously outdated over the last thirty years. Many of the species are also rather lumpy-looking and unappealing, and some are just direct stand-ins for recent mammals. For example, Australia is home to both kangaroo-like and koala-like dinosaurs for no apparent reason other than Australia just being Like That, and South America features very specific anteater, manatee, glyptodont, and sabertooth predator analogues.
Then came Man After Man in 1990. Initially intended to be a direct sequel to After Man, Dixon’s original concept would have shown the impact of humans arriving in the Posthomic period, escaping their dying overpopulated civilization by time-traveling into the far future and promptly causing ecological devastation and a mass extinction all over again.
But that was not to be.
Instead the publisher pushed Man After Man in a very different direction, and it became an “anthropology of the future” chronicling the evolution of post-human species genetically engineered to adapt to climate change. Rather grotesque and full of dystopian nightmare fuel, even Dixon himself has called it “a disaster of a project”.
(It also suffers from what may be plagiarism, with some of the designs looking worryingly close to those of another project that had been pitched several years previously by Wayne Barlowe. It remains unclear how much involvement and control Dixon actually had with Man After Man after the drastic change in concept, or whether he was even aware of this similarity.)
Some of Dixon’s works became especially popular in Japan, with After Man being adapted into both a stop-motion documentary and an animated short, and even having merchandise, and a manga version of The New Dinosaurs.
Later Dixon was a consultant and creature designer on the 2002 documentary series The Future Is Wild, a sort of “spiritual successor” to After Man – although due to the rights to After Man being held by Dreamworks at the time, none of his previous creations could be used. Depicting various ecosystems 5 million, 100 million, and 200 million years in the future, this series featured memorable speculative species such as whale-like gannets, giant carnivorous bats, sauropod-sized tortoises, four-winged birds, aerial fish, and arboreal squid.
The Future Is Wild was also adapted into both a manga and a Canadian-produced animated children’s series. There were plans over the years for a sequel documentary series, a feature film version, and then more recently a full reboot (including that VR game with the questionably-designed titan dolphin), but all seem to have succumbed to development hell.
Eventually Dixon’s original premise for Man After Man was reworked into Greenworld in 2010 (so far only ever published in Japanese), using his own artwork and changing the setting to an Earth-like exoplanet populated by animals descended from radially-symmetrical starfish-like ancestors. A generation ship carrying thousands of humans fleeing a devastated Earth arrive and colonize the planet – and over the next few centuries repeat history by exploiting the native lifeforms and putting Greenworld through multiple ecological catastrophes, causing a mass extinction that leaves the entire biosphere in ruins.
At some point another project called Microplatia was also developed for a planned exhibition on alien life at the Science Museum in London, but due to sponsorship issues was never completed.
Tomorrow: an expedition to a different alien world.
Spectember #09: Wading Hesperornitheans

Transcript for the text on the image under the cut:
Continue reading “Spectember #09: Wading Hesperornitheans”Spectember #08: Giant Cavy-Ungulates

Transcript for the text on the image under the cut:
Continue reading “Spectember #08: Giant Cavy-Ungulates”Spectember #07: Arboreal Crawfish

Transcript for the text on the image under the cut:
Continue reading “Spectember #07: Arboreal Crawfish”Spectember #06: Aquatic Afrosoricidans

(The submitted prompt here was “some kind of fully aquatic non cetacean or dugong mammal with a different bodyplan”.)
Transcript for the text on the image under the cut:
Continue reading “Spectember #06: Aquatic Afrosoricidans”Spectember #05: Frilled Fliers

{kind=link}
Transcript for the text on the image under the cut:
Continue reading “Spectember #05: Frilled Fliers”