Globicetus hiberus, a 5m long (16′4″) beaked whale from the Atlantic coast of Portugal and Spain. Its fossils can’t be easily dated since they were fished up from the seafloor, but it was probably around Early-to-Mid Miocene in age (~20-14 mya).
Its skull sported an odd bony sphere at the base of its snout, just in front of the melon, which appears to have been larger and more prominent in males than in females. Many modern beaked whales also have sexually dimorphic crests, ridges, and domes in their skulls, and these structures may function as sort of “internal antlers” – a display structure the whales can “see” via echolocation that signals their size, strength, and health to each other.
Let’s finish off this month the same way we started: with flying vertebrates without any transitional forms!
Much like the pterosaurs, bats appear suddenly in the fossil record already fully flight-adapted. Despite being the second-largest group of mammals, bats’ small fragile bones and terrestrial habitats make fossils of them incredibly rare, and transitional forms are still entirely unknown. (Even the ancestral form illustrated above is a generic hypothetical mammal!)
The most “primitive” known bats come from the Early Eocene* (~55-52 mya) and various early representatives have been found as far apart as North America, Europe, India, and Australia – indicating they were already a widespread and diverse group by that time, and making it difficult to pin down just where and when they actually might have originated.
*I’ve seen mentions of a potential bat-like tooth from the Late Cretaceous of South America, but can’t find any actual references for it. So it’s possible bats may even have evolved before the K-Pg extinction.
Although bats were once thought to be related to archontans (treeshrews, colugos, and primates) based on morphological similarities, more recent genetic studies have shown them to instead be grouped with the laurasiatheres (eulipotyphylans, carnivorans, pangolins, ungulates, and whales). Based on this phylogenetic position the earliest ancestors of bats may have been small tree-climbing shrew-like animals who evolved flight while leaping in pursuit of insects. They might even be closely related to an obscure group called nyctitheriids – but without a lucky find of an exceptional fossil, we just don’t know.
Based on the measurements of the teeth it was probably one of the larger eutriconodonts, close in size to Jugulator. I can’t find any body size estimates, but it may have have a total length of around 50-60cm (20-24″).
Plenty of fossil mammals are known solely from teeth, but what’s most interesting about this one is that its remains were found in coastal marine deposits without any signs of degradation or transport damage by water currents. This indicates the animal probably died at sea very close to the location where it was preserved.
A fewother eutriconodonts are now known to have been semi-aquatic, so Ichthyocondon might have been adapted to a similar lifestyle, making it one of the earliest known marine mammals. Another potential explanation is that it was a Volaticotherium-like glider that got blown out to sea.
As with many of this month’s paleontological mysteries, we need some more substantial fossil remains to know for sure. I’ve reconstructed it here as both main possibilities, as an otter-like semi-aquatic animal and as a patagium-bearing glider.
Almost a century later that one skull is still all we have. And despite this animal’s popularity among paleo-fans, we actually know very little about it.
It was originally classified as a mesonychian, leading to the many many depictions of it as a sort of “big bad wolf”. But more recent studies have placed it in the even-toed ungulates instead, with some suggestions that it might be most closely related to entelodonts, hippos, and whales.
Although it was certainly a big animal, it may not have been the giant “super predator” it’s often depicted as – its teeth aren’t particularly specialized and resemble those of entelodonts, suggesting it may have been more of an opportunistic omnivore than a dedicated carnivore.
Without more material we just don’t know for certain. So, frustratingly, the rest of Andrewsarchus’ body remains a mystery.
I’ve reconstructed it here based on one of its more obscure possible relatives: the anthracotheres, a group which may have been closely related to modern hippos. Scaling its body proportions to these animals produces rough measurements of about 1.45m tall at the shoulder (4′9″) and 3m long (9′10″), or about the same size as some of the big entelodonts or large modern bears.
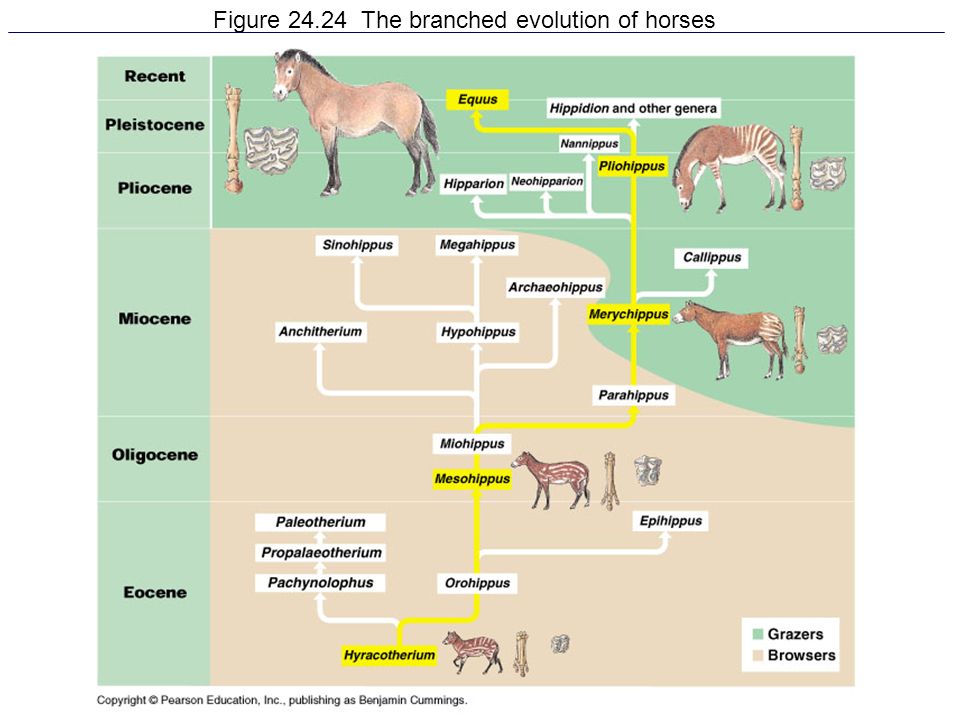
Horse evolution is often represented as a simple progression from Eohippus* to modern Equus, but it was actually a lot more complicated than that – and some ancient horses had some very odd things going with their snouts…
(* For a long time Eohippus was considered synonymous with Hyracotherium, but more recently has been split back off as its own genus again.)
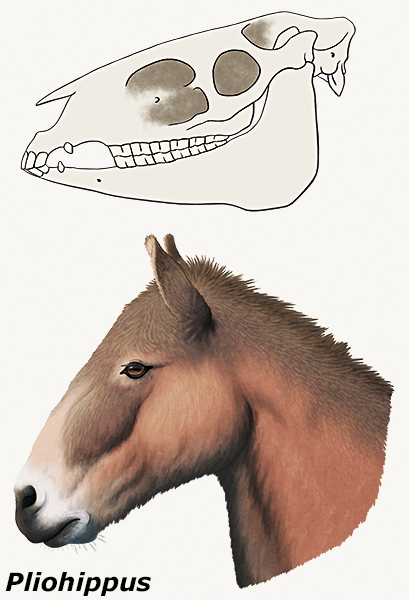
Pliohippus sp. skull and head reconstruction
Pliohippus, from the Middle Miocene of North America (~15-12 mya), and several of its other close relatives had especially large, deep recesses in their skulls, usually referred to as “preorbital fossae”.
And the purpose of these holes is still unknown. Although superficially similar depressions are seen in various other perissodactyl groups, they vary in position and structure and probably weren’t all homologous.
Ideas have included resonating chambers, some sort of glands, inflatable sacs, or attachment sites for complex lip musculature.
Hippidion sp. skull and head reconstruction
Meanwhile Hippidion from the Pleistocene of South America (2 million – 10,000 years ago) had especially long and domed nasal bones. This must have supported an enormous nasal area – possibly giving it a saiga-like air-conditioning system, a highly sensitive sense of smell, or perhaps even some sort of prehensile proboscis-like snout.
Unless we find some exceptional soft-tissue preservation, the facial anatomy of these equids is going to remain enigmatic.
Homalodotherium, a South American notoungulate mammal from the Early-to-Middle Miocene of Patagonia (~20-15 mya). Standing about 1.4m tall at the shoulder (4′7″), it seems to have convergently evolved to fill the same selective browsing niche as the North American chalicotheres and the later giant ground sloths.
Despite being an ungulate it had claws rather than hooves, and walked plantigrade on its hind feet but digitigrade on its front feet. It would have been capable of rearing up bipedally to pull down branches with its long forelimbs, with the shape of its nasal bones suggesting it may have also had a prehensile upper lip to help it strip off vegetation while feeding.
Lycopsis longirostrus*, a sparassodont mammal from the Middle Miocene of Colombia, South America (~14-12 mya). About 35cm tall at the shoulder (1′2″), it was originally thought to be a dog-like or thylacine-like animal, but studies of a well-preserved specimen have shown that it had an unusual mix of anatomic features in its limb bones, with adaptations for both climbing and running.
It was probably a terrestrial ambush hunter, preying on rodents and other small animals, but also had the ability to grasp and climb trees in its densely forested environment – especially useful for avoiding the land crocodiles that shared its habitat.








{kind=link}
_(18159159682).jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}