

Spinosaurid teeth were first found in the 1820s in England, but were misidentified as belonging to crocodilians. It wasn’t until nearly a century later that Spinosaurus aegyptiacus was discovered and recognized as a dinosaur – and it would be another century after that before we really started to learn anything about it.
1910s
The first fossils of Spinosaurus were discovered in Egypt in the 1910s. With only a few fragments of its skeleton known it was an enigma right from the start, hinting at a large and very strange theropod dinosaur with crocodile-like teeth, an oddly-shaped lower jaw, and elongated neural spines on its vertebrae that seemed to be part of a huge sail.
A few possible extra fragments were found in the 1930s, but overall these few pieces were all that was known of Spinosaurus for a long time.
The fossils were kept in the Paleontological Museum in Munich, Germany,a building that was severely damaged during a bombing raid in World War II. Many important specimens were destroyed, including Spinosaurus, and only the published drawings and descriptions of the bones remained.
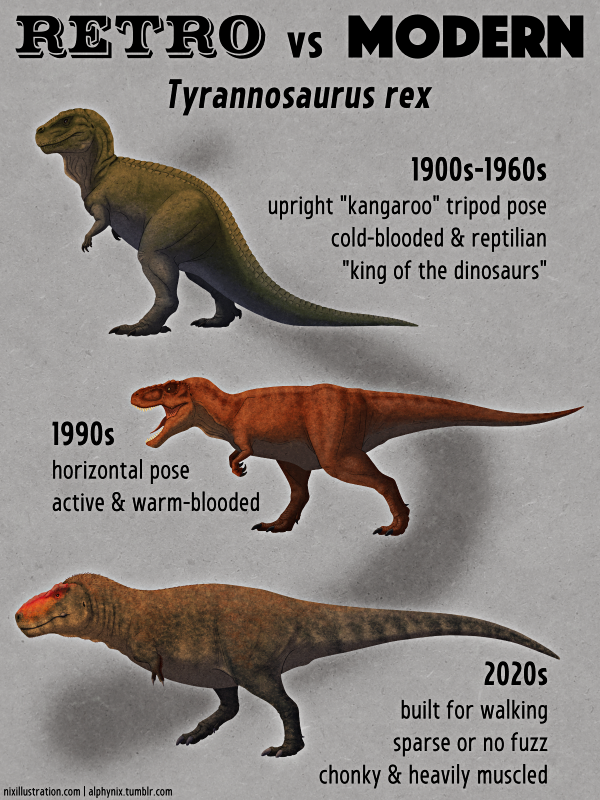
So for the next several decades Spinosaurus remained a very poorly-understood mystery. During this period it was generally depicted as a generic “carnosaur“, often modeled on something like Megalosaurus, in the standard-for-the-time tripod pose and with a Dimetrodon-like sail on its back.
Interestingly a 1930s skeletal reconstruction shows Spinosaurus with an unusually long torso and fairly short legs, details that are surprisingly modern despite the retro posture.
1990s
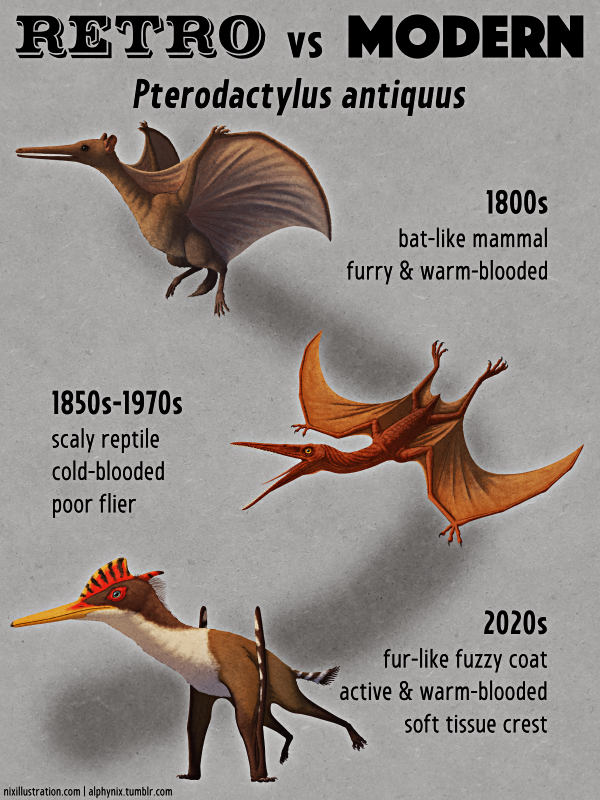
In the 1980s some partial snout bones from Niger were recognized as having similarities with the jaw of Spinosaurus. Around the same time the fairly complete skeleton of Baryonyx was discovered, and along with further spinosaurid discoveries in the mid-to-late 1990s a decent idea of what Spinosaurus might have looked like began to emerge.
It was reconstructed with a long kinked crocodilian-like snout, a ridged bony crest in front of its eyes, an S-curved neck, and large thumb claws on its hands – an interpretation that was heavily popularized by Jurassic Park III in the early 2000s, bringing this enigmatic dinosaur to public attention and portraying it as a fearsome super-predator bigger than Tyrannosaurus.
2020s
Despite attempts to locate more complete Spinosaurus remains, only fragments continued to be found, and it remained a frustratingly poorly-known species even into the early 2010s.
Finally, in 2014, almost a full century after it was first described and named, Spinosaurus started to reveal its secrets with the announcement of the discovery of the most complete skeleton so far, discovered in the Kem Kem fossil beds in Morocco. Its body was still only partially represented, but it included skull fragments, part of a hand, a complete leg and pelvis, some sail spines, and several vertebrae from the neck, back, and tail.
And nobody was expecting what these pieces revealed.
It had a very long torso and proportionally short stumpy legs, and was reconstructed with a huge distinctive “M-shaped” sail on its back. Its feet had flat-bottomed claws and its “dewclaw” toe was enlarged into an extra weight-bearing digit – adaptations for spreading its weight over soft muddy ground, and suggesting its feet may also have been webbed. Initially it was also presented as possibly being quadrupedal, due to how far forward its center of mass seemed to be, reviving an odd idea from the late 20th century.
Along with its long crocodile-like head and conical teeth, this was interpreted as evidence it was a semiaquatic fish-eating swimming animal – potentially making it the first known semiaquatic non-avian dinosaur. Spinosaurids had been suggested to be specialized piscivores before, especially since Baryonyx had been found with fish scales in its stomach, but they were generally assumed to be more like modern grizzly bears, wading into water to hunt but not being habitual swimmers. Spinosaurus’ weird croco-duck proportions, however, seemed like they might be much more suited to watery habitats than to the land.
Since Spinosaurus had become a popular dinosaur with the general public by that point, the discovery was big news – and a big controversy for a while. It was so bizarre that some paleontologists were skeptical of the radical new interpretation, wondering if the measurements of the skeleton were correct or if the short legs were even from the same individual or the same species as the rest of the bones.
After a while the new proportions were accepted as fairly accurate, and over the next few years attention turned to instead figuring out just how this animal worked and how aquatic it actually was. An earlier isotope analysis of its teeth supported a semiaquatic lifestyle similar to crocodiles and turtles, but a buoyancy study argued that it might not have been able to dive below the water suface and its sail made floating unstable – but also found that its center of mass was closer to its hips than previously calculated, suggesting it could walk bipedally after all.
Then in 2020 came another surprise: more of the tail of the new specimen had been found, and it was just as weird as the rest of Spinosaurus. Its tail was a huge vertically flattened paddle-like fin supported by long thin neural spines and chevrons, resembling a giant eel or newt more than a dinosaur and also giving some more weight to the idea that it was a swimmer.
Our modern view of Spinosaurus is still evolving, and likely to be full of even more surprises in the future as we discover more about this unique dinosaur. But we at least know it lived in what is now North Africa during the Late Cretaceous, about 99-93 million years ago, and whether it was a swimmer or wading generalist predator it was one of the largest known theropods to ever live, estimated to have reached around 16m long (~52ft).
While the “M-shaped” sail reconstruction has been popularized by the recent discoveries, the exact shape of this structure is still unknown. Like with other sailbacked animals it’s also not clear what it was for, with ideas including temperature regulation, visual display, supporting a fatty hump, and a potential hydrodynamic adaptation.
EDIT: And while I was working on this entry (late March 2022) I missed that another study had just come out with more anatomical support for swimming Spinosaurus!







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}