
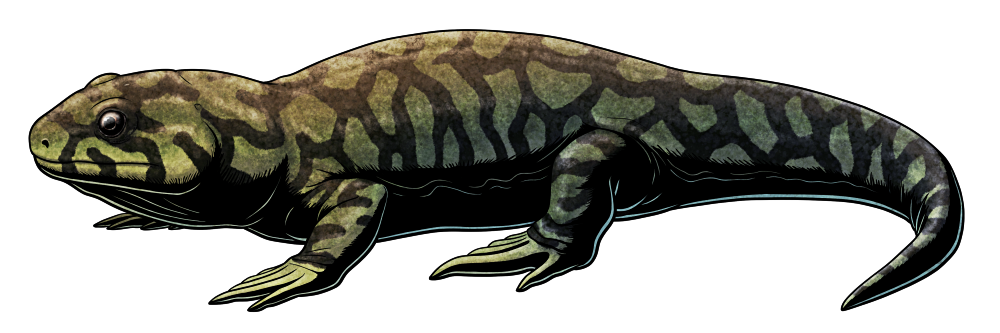
During the Early Carboniferous, around 330 million years ago, the region that is now the East Kirkton Quarry in Scotland was located close to the equator, with a lush tropical climate and volcanic hot springs dotting the landscape. It preserves fossils of some of the earliest known fully terrestrial tetrapods, and a recent discovery shows how some of these animals were already experimenting with the shapes of their feet to better get around on land.
Termonerpeton makrydactylus is only known from a partial skeleton, and shows a mix of anatomical features that make identifying its exact evolutionary relationships rather difficult – but it was probably a very early reptilomorph, closer related to amniotes than to lissamphibians. It may also have been very closely related to the equally enigmatic Eldeceeon and Silvanerpeton from the same region, but was almsot twice their size with a estimated total length of around 70cm (2’4″).
It would have resembled a rather heavily-built lizard-like or salamander-like animal, with fairly stumpy legs and probably lacking claws on its digits. While it would have had spindle-shaped scales on its underside, and possibly small rounded scales along its sides and back, these were bony structures embedded in its skin and probably weren’t very visible externally in life.
But Termonerpeton‘s most surprising feature was its proportionally large feet with especially elongated fourth toes, which would have helped to extend its stride length for energy-efficient terrestrial locomotion and to stabilize its movement on unstable surfaces – a much more “advanced” amniote-like arrangement than expected in such an early reptilomorph, and convergently similar to to the foot shapes of some modern lizards. Its fourth toe was also unusually chunky, suggesting it may even have been bearing most of its weight on just that one digit when walking.








{kind=link}