(This is a couple of days late for Halloween, but since this October saw the description of a new dinosaur species with a particularly spooky name, I couldn’t resist putting it into the schedule anyway.)
Spectrovenator ragei was an early member of the abelisaurid lineage, living in southeastern Brazil during the Early Cretaceous, about 120 million years ago. It was one of the smallest known abelisaurids, measuring just 2m long (6’6″), and lacked a lot of the skull specializations seen in larger-bodied Late Cretaceous forms like Carnotaurus, suggesting it was more of a generalist predator.
Its genus name translates to “ghost hunter” due to it being found underneath the fossil remains of another dinosaur entirely – a “ghost” unexpectedly appearing when the specimen was being prepared – but it’s extra appropriate since it also helps to fill in a rather sizeable ghost lineage in the fossil record of abelisaurids.
Named after the mythological bird-like Anzû – and also nicknamed “the chicken from hell” – Anzu wyliei was one of the larger known oviraptorosaurs, measuring about 3m long (9’10”).
Its fossils are some of the most complete for a North American member of this dinosaur group, with four different specimens representing about 80% of the whole skeleton.
Living right at the end of the Cretaceous, about 66 million years ago in North Dakota and South Dakota, USA, Anzu inhabited the ancient floodplains of Hell Creek and appears to have been a fairly fast-moving omnivorous generalist. It had a large crest on its head made of rather fragile thin-walled bone, which may have been used for display or sound amplification similar to the casque of modern cassowaries.
Some of the fossil specimens also show evidence of healed injuries, including a broken rib and an arthritic toe.
Many modern predatory birds have enlarged claws on their second toes, similar to those of their paravian dinosaur ancestors – with seriemas being a particularly good example.
Seriemas are part of a lineage known as cariamiformes, highly terrestrial birds that were widespread across most of the world but are today represented today by only two living species in South America. During the Cenozoic this group repeatedly evolved into large predatory flightless forms like the the phorusrhacids and bathornithids, and were probably the closest avians ever got to recreating the “carnivorous theropod” body plan and ecological niche.
And yet none of them ever seem to have experimented with more dromaeosaurid-like claws.
…With one known exception.
Qianshanornis rapax here lived in East China during the mid-Paleocene, about 63 million years ago. It was a small cariamiform, probably around 30cm tall (1″), and is only known from fragmentary fossil material – but part of those fragments was a fairly well-preserved foot. And the bones of its second toe were unlike any other known Cenozoic bird, shaped incredibly similarly to those of dromaeosaurids and suggesting it may have had the same sort of big hyperextendible “sickle claw”.
While it had sturdy legs and short wings, and probably spent a lot of time walking on the ground like other cariamiformes, it was probably also still a fairly strong flier based on the known anatomy of its arms and shoulders.
Unfortunately, though, its head and claws were entirely missing, so without more fossil discoveries it’s hard to say anything definite about its ecology. I’ve restored it here based on other predatory cariamiformes, but since it was also closely related to a herbivorous species it’s not clear whether Qianshanornis was truly a dromaeosaur-mimic or if something else was going on with that unique second toe.
They were fairly small stocky quadrupedal dinosaurs, sort of pig-like, with short deep jaws and powerful beaks adapted for eating fibrous low-level plants like ferns and cycads – and to process such tough food they even evolved a chewing style similar to mammals like rodents.
Prenoceratops pieganensis here is known from the Two Medicine Formation bone beds in Montana, USA, dating to about 74 million years ago. Around 1.5-2m long (~5′-6’6″), it was very similar to its later relative Leptoceratops, but had a slightly lower, more sloping shape to its skull.
The pelagornithids, or “pseudotooth birds”, were a group of large seabirds that were found around the world for almost the entire Cenozoic, existing for at least 60 million years and only going completely extinct just 2.5 million years ago.
Their evolutionary relationships are uncertain and in the past they’ve been considered as relatives of pelicaniformes, albatrosses and petrels, or storks, but more recently they’ve been proposed to have been closer related to ducks and geese instead.
Whatever they were, they were some of the largest birds to ever fly, and many of the “smaller” species still had wingspans comparable to the largest modern flying birds.
But their most notable feature was their beaks. Although at first glance they look like they were lined with pointy teeth, these structures were actually outgrowths of their jaw bones covered with keratinous beak tissue. While these bony spikes would have been useful for holding onto slippery aquatic animals like fish and squid, they were actually hollow and relatively fragile so pelagornithids must have mainly caught smaller prey that couldn’t thrash around hard enough to break anything.
The serrations also only developed towards full maturity, and the “toothless” juveniles may have had a completely different ecology to adults.
Pelagornis chilensis here was one of the larger species of pelagornithid, with a wingspan of 5-6m (16’4″-19’8″), known from the western and northern coasts of South America during the late Miocene about 11-5 million years ago.
Like other pelagornithids it was highly adapted for albatross-like dynamic soaring, with long narrow wings that allowed it to travel huge distances while expending very little energy – but with its proportionally short legs it would have been clumsy on the ground and probably spent the vast majority of its life on the wing, only returning to land to breed.
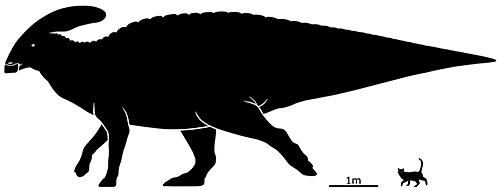
But by far the most recognizable of the crested hadrosaurs is Parasaurolophus walkeri, with its long curved backwards-pointing tubular crest.
This particular species was mid-sized for the genus, growing up to around 10m long (32’10”) and is known from Western North America during the Late Cretaceous, about 76-73 million years ago.
Its crest was intermediate in size and shape between the other two known species. The larger Parasaurolophus tubicen had a longer and slightly straighter crest, while the smaller Parasaurolophus cyrtocristatus had a shorter more strongly curved one. Juveniles developed these crests as they matured, starting off with much smaller bumps on their snouts that gradually grew backwards and upwards.
Some hadrosaur crests were purely for visual display, but in the lambeosaurine lineage that Parasaurolophus belonged to they also incorporated complex looping nasal passages that were probably used as resonating chambers, allowing each species to make a unique-sounding loud bellowing call to communicate with each other.
There are also rumors of a currently-undescribed specimen of Parasaurolpphus that has preserved soft tissue around its crest, possibly a keratinous covering or skin flaps that made it appear even larger and more flamboyant in life than the underlying bone. So I’ve given this reconstruction a speculative structure like that, along with hoof-like claws on its hands similar to those recently revealed for Edmontosaurus.
Much like their frilled relatives they had beaks at the tips of their snouts and large gut cavities for digesting plant matter, but they also had surprisingly sharp theropod-like teeth in front of their more standard herbivore teeth further back – suggesting they may also have been opportunistic omnivores, occasionally snacking on carrion or small animals similarly to modern pigs or bears.
Their striking-looking dome heads were probably used for combat, headbutting or flank-butting each other, and many fossil skulls show evidence of injuries that would have been caused by that sort of behavior.
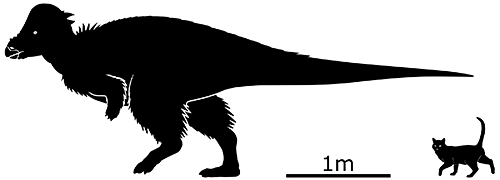
The eponymous Pachycephalosaurus wyomingensis lived in North America right at the end of the Cretaceous, about 70-66 million years ago. It was one of the largest of its kind, reaching lengths of around 4.5m (14’9″), and was characterized by a large bony dome-head surrounded by small blunt spikes.
But it turns out that was probably only what it looked like as a fully mature adult.
Recent discoveries of juvenile Pachycephalosaurus skulls confirmed a hypothesis proposed a few years earlier: these dinosaurs changed appearance drastically as they grew up, and younger individuals had been mistaken for separate species. They started off with domeless flat heads, bristling with long spikes (a form previously named Dracorex hogwartsia) then as they matured their domes began to grow (previously Stygimoloch spinifer) and by full maturity they had big domes with the spikes shrunk down to smaller stubbier knobs (the classic Pachycephalosaurus look).
This particular reconstruction depicts a Stygimoloch-like subadult individual, not quite fully mature and still sporting some longer spikes.
We can’t go through this month without having an appearance from the most famous group of weird-headed dinosaurs: the ceratopsids!
Their distinctive-looking skulls were highly modified from those of their ancestors, with large bony frills extending from the back of their heads, various elaborate horns and spikes, enormous nasal cavities, large hooked beaks at the front of their snouts, and rows of slicing teeth further back.
And while typically depicted as purely herbivorous, ceratopsids’ powerful parrot-like beaks and lack of grinding teeth suggest they may actually have been somewhat more omnivorous – the Cretaceous equivalent of pigs – still feeding mainly on plant matter but also munching on carrion and opportunistically eating smaller animals when they got the chance.
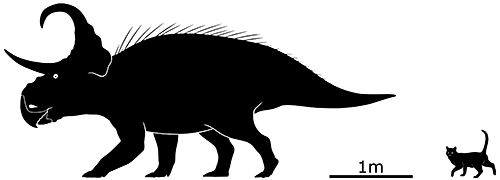
Machairoceratops cronusi here lived during the late Cretaceous of Utah, USA, about 77 million years ago. Only one partial skull has ever been found belonging to an individual about 4.5m long (14’9″), but it wasn’t fully grown and so probably reached slightly larger sizes.
It had two long spikes at the top of its frill, similar to its close relative Diabloceratops but curving dramatically forward and downwards above its face. Whether they were purely for display or used in horn-locking shoving matches is unknown, but either way it was a unique arrangement compared to all other known ceratopsids.
Dilophosaurus wetherilli is a fairly recognizable dinosaur thanks to its memorable appearance in the Jurassic Park franchise – but unfortunately that also means the popular image of it is completely wrong.
Rather than a small frill-necked venom-spitting creature, this early theropod was actually rather large, reaching around 7m long (~23′), and along with its distinctive double crests it also had a narrow snout with large teeth and a distinctive notch at the front of its lower jaw.
It lived in North America during the early Jurassic, about 196-183 million years ago, and while it wasn’t venomous its notched jaws were probably capable of delivering powerful bites to small struggling prey, much like the similar-looking ornithosuchids in the Triassic. Some structural similarities to the skulls of spinosaurids suggest it may have primarily eaten fish.
Its two bony crests were probably used for visual display, with juveniles only having small crests that fully developed as they matured. They also may have had a more extensive keratinous covering, so it’s not clear what their actual shape and full extent was in life.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}