It was a fairly small ornithomimid at around 2.5-3m long (~8-10′), and its leg proportions suggest it was less specialized for fast running than some of its other relatives. But its most unique feature was its hands, with extremely elongated metacarpal bones – giving it palms that were longer than its upper arm!
It’s unclear what the function of these unusual limbs was. Possibly they gave Mexidracon extra reach to hook its claws onto foliage and pull it within reach of its head, in a similar manner to ground sloths, chalicotheres, and therizinosaurs.
It was a member of a diverse group of Mesozoic birds known as enantiornitheans, which retained claws on their wings and often still had toothed snouts instead of beaks – and being part of the avisaurid family it was also one of the larger known examples of these birds, similar in size to a modern hawk at around 60cm long (~2′).
Although this species is only known from isolated foot bones, the remains have distinct enough anatomical features to show that Avisaurus had powerful gripping talons similar to those of modern hawks and owls, suggesting it had a similar lifestyle hunting small vertebrate prey in the ancient swampy Hell Creek ecosystem.
First discovered in the 1850s, Compsognathus longipes was the first theropod dinosaur known from a fairly complete skeleton, and also the smallest known non-avian dinosaur for over 130 years.
(A second specimen was also, briefly, the “first” aquatic non-avian dinosaur, but that’s another story.)
Living in what is now Europe during the late Jurassic, about 150 million years ago, it was a lightly built animal with long legs and a long tail, growing to around 1.2m long (~4′). Its hands seem to have had only two functional fingers, with the third being vestigial and possibly not even having a claw.
Skin impressions from about a third of the way along its tail show small bumpy scales – but since other compsognathids like Sinosauropteryx are known to have been covered in fur-like feathers, this likely means that just that particular region of Compsognathus’ body wasn’t fluffy.
Some of Compsognathus‘ diet is known for certain, since preserved gut contents show it fed on smaller vertebrates like lizards and rhynchocephalians. The remains of a lizard in the stomach of one specimen were even identified as belonging to a previously-unknown species, Schoenesmahl dyspepsia, with the dismembered nature of the skeleton suggesting Compsognathus tore its prey into bite-sized chunks in a similar manner to modern predatory birds.
While birds are one of the few animal groups to have achieved powered flight, they’re also very prone to losing their aerial abilities. Many times over their evolutionary history, multiple different bird lineages have convergently become secondarily flightless – and Patagopteryx deferrariisi was one of the earliest known examples of this.
Living during the Late Cretaceous, about 86-84 million years ago, in what is now the northern part of ArgentinePatagonia in South America, Patagopteryx was roughly the size of a modern chicken at around 50cm long.
When it was first discovered it was classified as a ratite, but soon after it was recognized as actually being a much earlier type of bird, an early ornithuromorph only distantly related to any modern groups.
It had small wings, little-to-no keel, and no wishbone, indicating it lacked the large powerful musculature required for flight. Its legs were quite long, with large feet with all four toes facing forward – proportions that suggest it was built more for walking than for high-speed running.
Growth rings in its bones also show that it had a much slower growth rate than modern birds, taking several years to reach adult size.
I’m still trying to work through that big pile of speculative evolution concepts from a few years ago, so I’m hoping to make this month sort of a “lightning round” to finally clear out the backlog.
(I’m not going to set a definite posting schedule this year because things are pretty chaotic right now. But I’ll try to fit in as many as I can!)
So let’s start off with a concept from an anonymous submitter, who requested a “kiwi/sengi niche alverezsaur”:
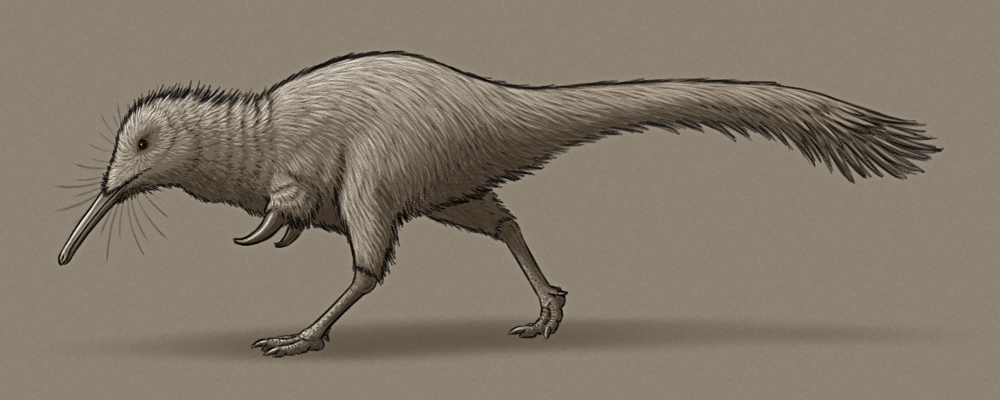
Khamartaia dolabella is similar in size and build to Shuvuuia, about 1m in length (3’3″), with slender legs and stumpy arms with massive thumb claws. Unlike its close relatives, however, it has small eyes and fairly poor vision, relying more on its other senses to forage around during the darkness of night.
It has an acute sense of smell, and its long narrow snout is full of highly touch-sensitive nerves, allowing it to probe around for invertebrate prey in soil, undergrowth, and cracks and crevices. Its chunky thumb claws are used to dig up burrows and to tear through bark to access deeper insect nests.
It mainly relies on its long legs to sprint away from threats, although with its poor eyesight these escapes are often rather ungainly.
The next part of the Crystal Palace Dinosaur trail depicts the Jurassic and Cretaceous periods. Most of the featured animals here are actually marine reptiles, but a few dinosaur species do make an appearance towards the end of this section.
Head, flipper, and tail details of the Temnodontosaurus. A second ichthyosaur is just barely visible in the background.
Ichthyosaurs were already known from some very complete and well-preserved fossils in the 1850s, so a lot of the anatomy here still holds up fairly well even 170 years later. They even have an attempt at a tail fin despite no impressions of such a structure having been discovered yet! Some details are still noticeably wrong compared to modern knowledge, though, such as the unusual amount of shrinkwrapping on the sclerotic rings of the eyes and the bones of the flippers.
The semi-aquatic spinosaurids were an unusual exception to this with six or seven teeth per premaxilla – and one particular member of this lineage seems to have been just a little bit weirder.
Baryonyx walkeri lived during the early Cretaceous, around 130-125 million years ago, in what is now southeast England. About 9m long (~30′), it had distinctive enlarged curving claws on the first fingers of its hands, along with a long narrow snout with a “rosette” at the tip followed by a notch (a shape convergent with the jaws of modern pike conger eels).
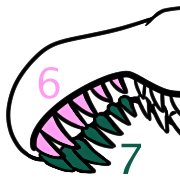
And that premaxillary rosette had a strangely asymmetrical arrangement of teeth.
The left side had six teeth, and the right side had seven.
Why? We don’t know!
Baryonyx skull material is rare and fragmentary, so it’s unclear if this was actually a characteristic feature of the species or if the known asymmetric rosette just represents an unusual individual.
Halszkaraptorines were a group of small dromaeosaurids known only from the Late Cretaceous of Mongolia. They were odd little raptors with flattened snouts, long necks, and flipper-like arms – features that suggest they were specialized for swimming, making them the second known lineage of semi-aquatic non-avian dinosaurs after the spinosaurids.
Natovenator polydontus lived in what is now the Gobi Desert in southern Mongolia, around 72 million years ago. The size of a small duck, about 45cm long (18″), it had jaws full of many needle-like teeth, a long flexible goose-like neck, and a streamlined body with a wide flattened ribcage convergently shaped like those of modern diving birds.
Although it had long strong legs, these don’t show much in the way of aquatic specializations and would have been used more for walking and running on land. Instead it may have used its flipper-like arms to propel itself through the water, like modern penguins or auks.
It probably had a lifestyle similar to modern mergansers, swimming and diving in lakes and rivers, and preying on fish, amphibians, and aquatic invertebrates.
The ostrich-like “bird-mimic” dinosaur Ornithomimus was named in 1890, based on some hand and foot bones from Late Cretaceous-aged fossil beds in Colorado, USA.
The first ornithomimid known to science, it was initially thought to be a ornithopod, but then a few years later more fossil material revealed it was actually a theropod – and then it spent some time classified as a “megalosaur” before ornithomimids were finally recognized as being coelurosaurs in the early 20th century.
And for nearly a century after its discovery it was treated as a wastebasket taxon for any similar-looking fossil material from North America and Asia, with around 17 different species named within the genus. One of these was split off into Struthiomimus in 1917, but it wasn’t until much later that the rest began to get sorted out.
A review of known Ornithomimus fossils in the early 1970s renamed a couple more species into the new genera Archaeornithomimus and Dromiceiomimus, and dismissed most of the remaining species as dubious or invalid. Just two valid species now remained: the original Ornithomimus velox from Colorado, and Ornithomimus edmontonicus from Alberta, Canada.
Ornithomimus edmontonicus
Since then opinions have gone back and forth about some of the other Ornithomimus species. For a while Dromiceiomimus was merged back into Ornithomimus, but more recently it’s been found to have distinct limb proportions and was probably actually a separate genus after all. Another species that’s usually considered to be part of Struthiomimus is also sometimes instead classified as an Ornithomimus instead.
Really all of the North American ornithomimids are in need of a modern taxonomic revision – especially since Ornithomimus edmontonicus shows enough anatomical variation that it might actually represent a species complex of multiple very similar forms, which might get split apart in the future if anyone can figure out how to reliably distinguish them.
But during the 1960s and 1970s this arrangement began to break down. A better understanding of groups like dromaeosaurs revealed a confusing mixture of traditional “carnosaur” and “coelurosaur” anatomical features, and paleontologists struggled to figure out where these sorts of theropods actually fit in.
The development of cladistic methods from the 1970s onwards led to efforts to clean up the coelurosaur wastebasket, trying to figure out a more accurate version of these animals’ evolutionary relationships. After briefly collapsing Coelurosauria down to just coelophysoids and “coelurids“, the growing recognition of modern birds as living theropod dinosaurs eventually resulted in the group being properly redefined in the 1980s as “birds, and all theropods closer related to them than to carnosaurs“.
Clockwise from the left (not to scale): Citipati osmolskae, Albertosaurus sarcophagus, Yi qi, Sinosauropteryx prima
The coelophysoids were finally removed entirely, reclassified as a much earlier branch of theropods – but quite a few of the other groups from earlier concepts of Coelurosauria survived this reshuffling, with the compsognathids, ornithomimids, oviraptorosaurs, dromaeosaurs, and troodontids all proving themselves to have really been closely related the whole time. Meanwhile the tyrannosauroids were brought back in, along with the therizinosaurs, alvarezsauroids, and a whole bunch of paravian and avialan lineages.
(Megaraptorans might belong somewhere in the coelurosaurs, too – possibly being tyrannosauroids – but their classification is currently being disputed.)



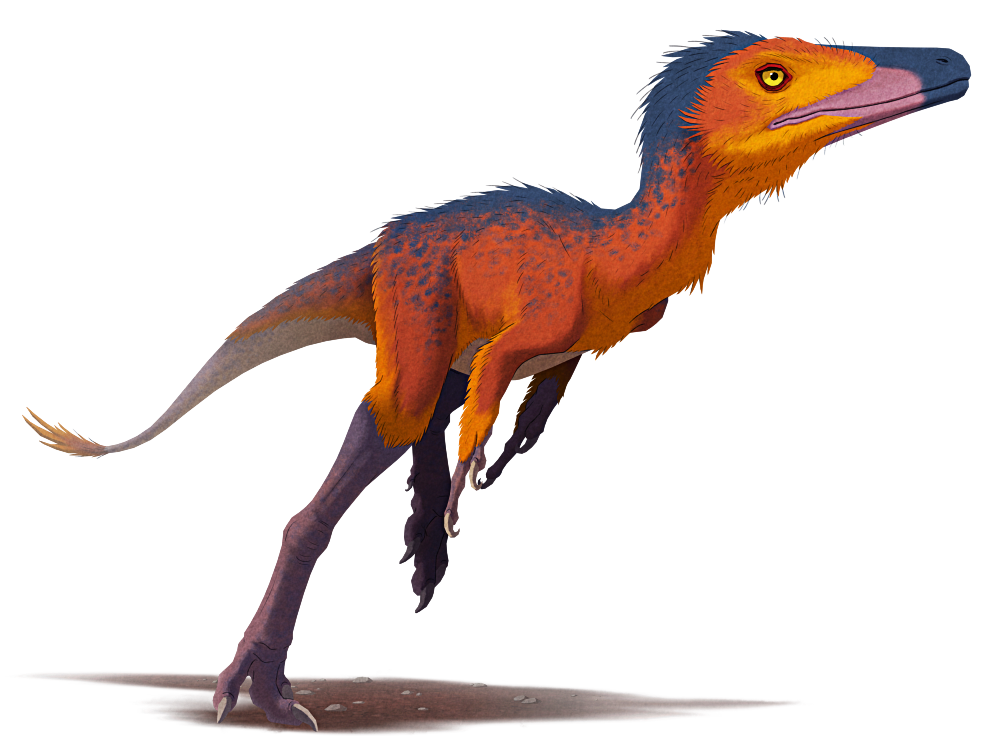









{kind=link}