

Along with its unique birds, Mauritius was also home to many endemic reptile species. In the absence of terrestrial mammals giant tortoises were the largest herbivores on the island, and various geckos, skinks, and snakes helped to fill out the rest of the vertebrate ecosystem.
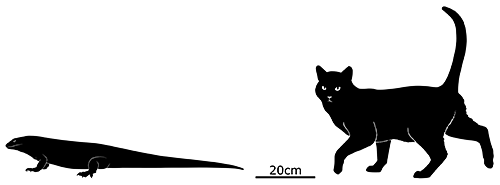
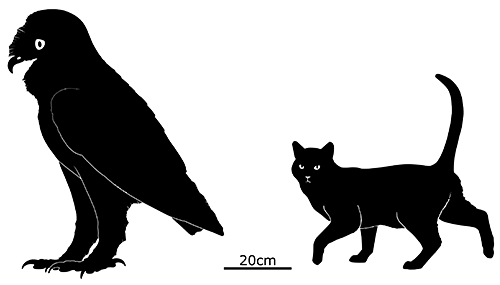
Leiolopisma mauritiana was a very large skink, one of the biggest ever known to have existed with a total length of around 80cm (2′7″). Its ancestors originated in Australasia, over 5600km away (~3500 miles) at least 3-4 million years ago – and they must have endured a particularly long ocean rafting journey without any island hopping stops, since none of the other islands along that route seem to have ever had populations of similar skinks.
It probably lived in rocky areas, possibly also being capable of digging burrows, and would have eaten an omnivorous diet of seeds, fruits, invertebrates, and smaller lizards and birds.
By the early 1600s it was already extinct, very soon after the arrival of humans, probably due to predation from invasive mammals like rats. However, its half-sized close relative Leiolopisma telfairii does still survive on rat-free Round Island a short distance to the north of Mauritius, and recent conservation efforts have been rebuilding its population and setting up new colonies on other nearby small islands.













.jpg){kind=link}
{kind=link}