
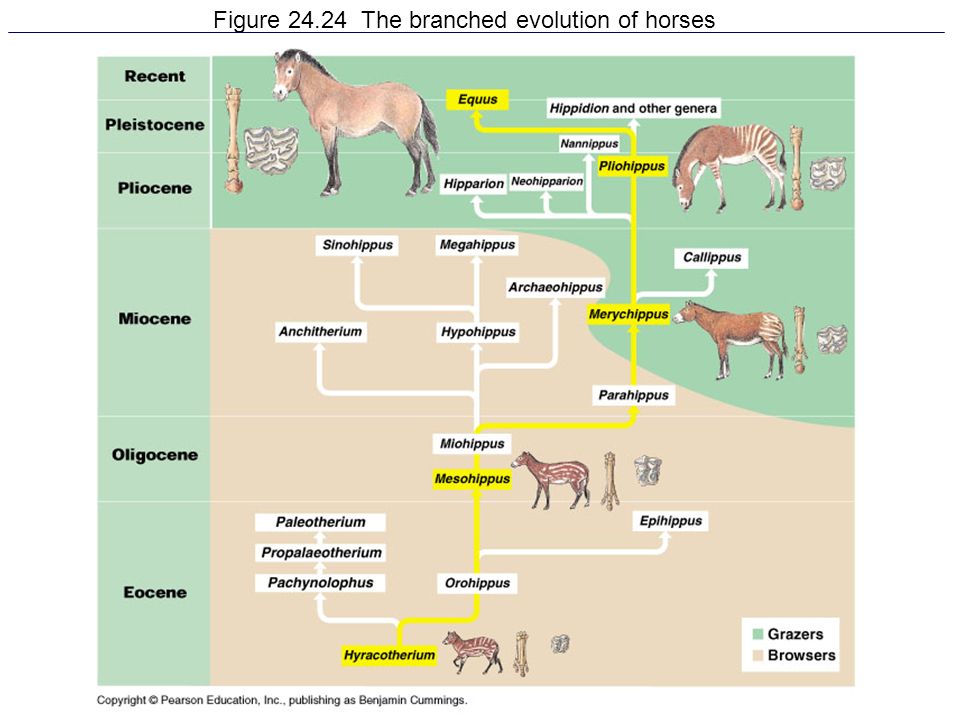
Horse evolution is often represented as a simple progression from Eohippus* to modern Equus, but it was actually a lot more complicated than that – and some ancient horses had some very odd things going with their snouts…
(* For a long time Eohippus was considered synonymous with Hyracotherium, but more recently has been split back off as its own genus again.)
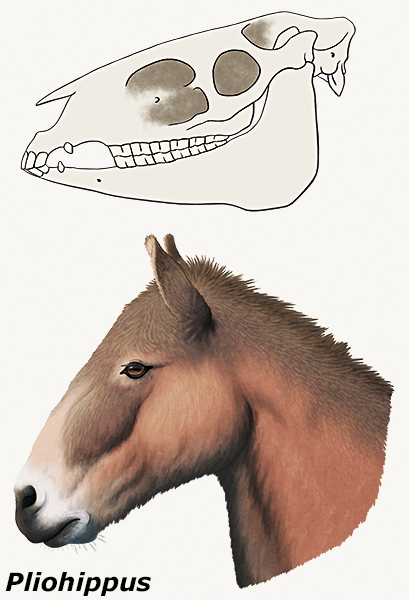
Pliohippus, from the Middle Miocene of North America (~15-12 mya), and several of its other close relatives had especially large, deep recesses in their skulls, usually referred to as “preorbital fossae”.
And the purpose of these holes is still unknown. Although superficially similar depressions are seen in various other perissodactyl groups, they vary in position and structure and probably weren’t all homologous.
Ideas have included resonating chambers, some sort of glands, inflatable sacs, or attachment sites for complex lip musculature.

Meanwhile Hippidion from the Pleistocene of South America (2 million – 10,000 years ago) had especially long and domed nasal bones. This must have supported an enormous nasal area – possibly giving it a saiga-like air-conditioning system, a highly sensitive sense of smell, or perhaps even some sort of prehensile proboscis-like snout.
Unless we find some exceptional soft-tissue preservation, the facial anatomy of these equids is going to remain enigmatic.










{kind=link}