

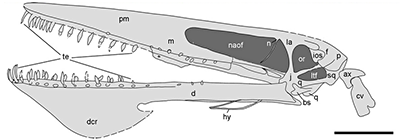
Longipteryx chaoyangensis, an enantiornithine from the Early Cretaceous of China, about 120 million years ago. With a body length of only around 15cm (6″), it had a long snout tipped with a few hooked teeth and feet capable of perching – features that indicate it may have lived very similarly to modern kingfishers, feeding on fish and small invertebrates in its swampy forest habitat.
The enantiornithines were a sort of “cousin” lineage to modern birds. Most had toothy jaws and clawed wings, and the wide variety in their skull shapes suggests that they were specialized for many different dietary niches. The entire group went extinct during the K-Pg mass extinction and left no living descendants, but during the Cretaceous they were the most widespread and diverse group of birds*, with fossils currently known from every continent except Antarctica.

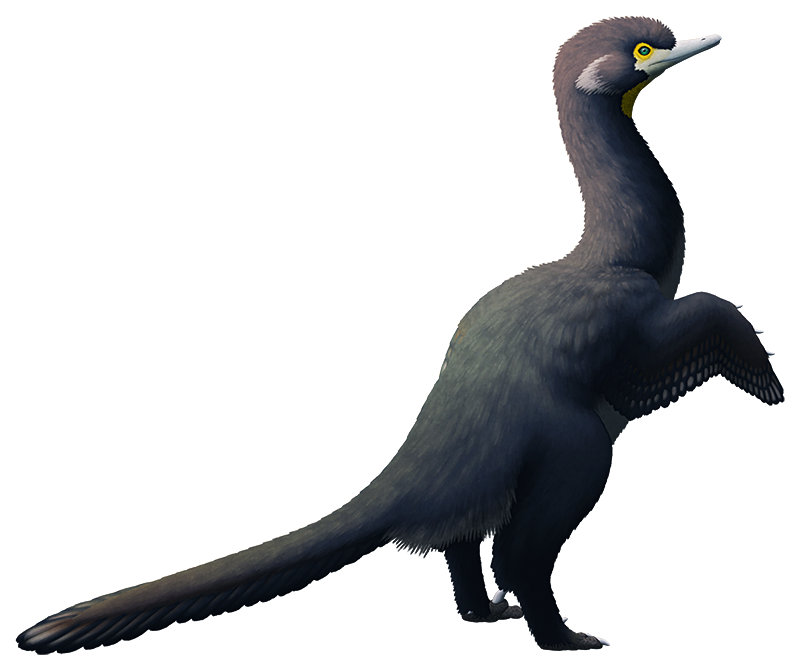








{kind=link}