It was rather small compared to its modern relatives, just 30-40cm in length (~1′-1’4″), with a long streamlined body armored with six rows of scutes. Its slender snout was lined with tiny teeth, and both its first dorsal fin and first anal fin were tall and elongated.
It probably wasn’t a very active swimmer, instead hovering near the surface and catching smaller prey with quick bursts of speed.
It lived around 50-48 million years ago during the early Eocene, in a shallow warm sea covering what is now Italy, with its three known fossil specimens all coming from the fish-rich Monte Bolca fossil beds.
About 60cm long (~2′), it had a round pancake-like body similar to many modern seafloor-dwelling stingrays – but uniquely it was also almost tailless, with only a tiny, slender, stingless tail.
It wasn’t a particularly strong swimmer, instead probably spending most of its time buried in the muddy seafloor sediment. When on the move it likely swam along just above the surface of the seafloor using undulations of its fins, foraging for smaller bottom-dwelling animals like worms, molluscs, crustaceans, and fish.
Living during the Late Devonian, about 375-359 million years ago, in subtropical waters covering parts of what are now North America and Europe, this species is known mostly just from the bony plates that covered its head and thorax. The rest of its skeleton was cartilaginous and rarely ever fossilized (only a few vertebrae and the pectoral fin are currently known), so its full body shape and size is poorly understood, and previous length estimates have ranged all the way up to 10m (33′).
…Except it turns out it wasn’t nearly that big.
Based on its head proportions, along with comparisons to more complete remains of other arthrodire placoderms, recent studies instead come up with a maximum length of about 4m (~13ft) – giving Dunkleosteus a much shorter-but-heavier chunky body shape, more like a tuna than a shark.
Preserved stomach contents in one fossil show remains of the fast-swimming cartilaginous fish Orodus – suggesting that much like the modern tuna it resembled, Dunkleosteus was also capable of bursts of high speed.
Ellimmichthyiformes were a group of ray-finned fish known from the early Cretaceous to the mid-Oligocene, about 140-30 million years ago. For much of that time they were quite widespread, found in various marine, estuarine, and freshwater environments across Africa, Eurasia, and the Americas.
Closely related to modern clupeiformes (herrings, sardines, and anchovies), and characterized by two rows of bony scutes – one in front of the dorsal fin and the other along the belly – they’re also known by the nickname “double‐armored herrings”.
Rhombichthys intoccabilis was a rather unusual-looking ellimmichthyiform from the mid-Cretaceous, around 95 million years ago. Living in shallow reef and lagoon waters covering what is now the West Bank in the Middle East, it was about 20cm long (~8″) and had a tall narrow dorsal fin along with incredibly elongated belly scutes that gave its body a rhombus-like profile.
Juveniles of this species seem to have lacked the extended belly scutes, instead having a much more rounded body shape. This may indicate that adults and juveniles occupied very different ecological roles, or that the distinctive scutes might have been a secondary sexual characteristic involved in displaying for courtship and reproduction.
Up to around 2.5m long (~8′), it’s known from several exceptionally well-preserved and near-complete skeletons.
It had a streamlined body with large pectoral fins, small pelvic fins, and a strongly keeled crescent-shaped tail fin. And although it was superficially shark-like in appearance, it was actually part of a lineage known as cladoselachids, which were much closer related to modern chimaeras than to sharks.
It’s unclear if Maghriboselache had two dorsal fins like its close relative Cladoselache, but some specimens preserve evidence of a chunky spine where the front dorsal fin would have been. Others show no sign of a front dorsal fin or spine at all, suggesting there may have been some sexual dimorphism going on in this species, with males having a spine (and possibly also an associated front dorsal fin) and females only having a rear dorsal fin.
But the most unusual feature of Maghriboselache was its nose.
It had a very broad snout with large and unusually widely-spaced nostrils, which would have given it the ability to “smell in stereo” and determine the direction of scents carried through the water much more precisely – making it the earliest known example of that sort of sensory specialization.
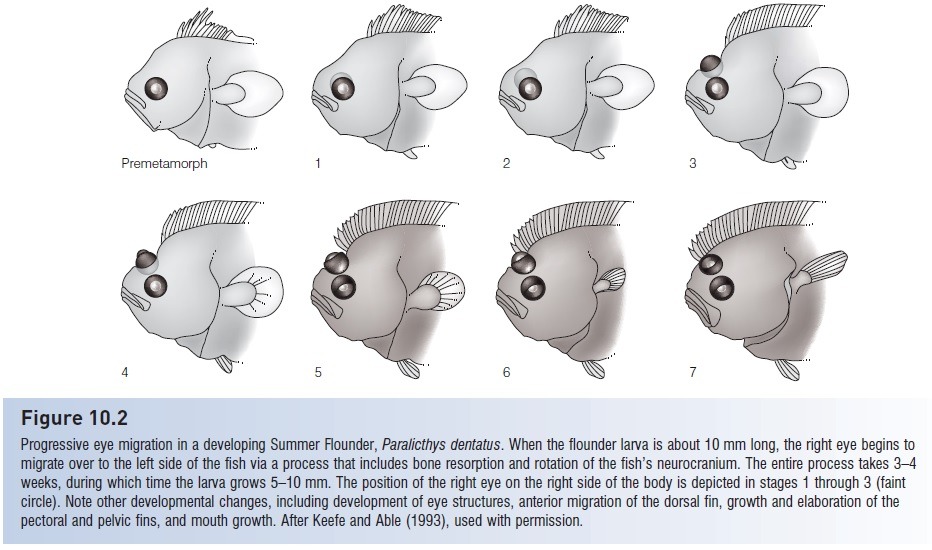
Modern flatfish are characterized by their highly asymmetrical skulls, with both eyes positioned on just one side of their head. They aren’t actually born this way, but instead they undergo “eye migration” as juveniles, twisting up their skulls to bring one eye across the top of the head.
Progressive eye migration in a developing Summer Flounder, Paralicthys dentatus. From Helfman et al (2009). The diversity of fishes. 2nd ed., Wiley-Blackwell.
This bizarre arrangement is the result of flatfish adapting to life laying flat on the seafloor, but instead of slowly widening and flattening themselves out they took an evolutionary “shortcut” by simply tipping their tall narrow bodies over onto one side. Initially this would have left one of their eyes unusable, but random mutations causing slightly asymmetrical skulls would have rapidly become highly advantageous to the earliest members of this lineage – and over time they just got wonkier and wonkier.
We’ve even found fossils of early flatfish in the “halfway there” stage of their evolution!
Amphistium paradoxum lived in what is now northern Italy during the Eocene, around 50-48 million years ago. About 20cm long (~8″), it had one eye partially migrated towards the top of its head, but not all the way around yet, showing a transitional state between its bilaterally symmetric ancestors and its more twisted-skulled modern relatives.
Unlike most modern flatfish Amphistium came in both “right-eyed” and “left-eyed” forms in equal numbers, suggesting that a genetic preference for a specific side also hadn’t developed yet.
This repeated “pristification” suggests that saws are just incredibly useful and relatively “easy to evolve” structures for these types of fish, being both highly sensitive to bioelectric fields and able to physically slash and stab to kill prey.
Onchopristis numida was a sawskate known from what is now Northern and Western Africa during the mid-Cretaceous, about 95 million years ago. Up to about 3m long (~10′), it lived in both saltwater and freshwater, and was probably a bottom-dwelling ambush predator similar to modern angelsharks.
Whenever a denticle was lost from its saw, a larger one would grow to replace it, and over the life of an Onchopristis this resulted in an increasingly extreme amount of saw asymmetry.
Modern pristified fish also have rather asymmetrical saws. Sawfish are commonly born with a different number of denticles on each side, while sawsharks add extra denticles of varying sizes as they age, with the ongoing replacement of lost denticles resulting in more uneven arrangements over their course of their lives.
It’s not clear if the asymmetry gives any sort of advantage to these fish – but if nothing else it probably doesn’t cause them any disadvantage, so there’s no evolutionary pressure to stay more symmetrical.
Most of them also had jaws full of round flat teeth used to crush hard-shelled prey, but some may instead have been herbivorous grazers similar to parrotfish.
And a couple of lineages even became carnivores.
Serrasalmimus secans lived in what is now Morocco during the late Paleocene, about 59 million years ago. Although only known from its jaws, the size of the fossil material suggests it was fairly large for a pycnodont, possibly around 80cm long (~2’8″).
It had sharp flesh-cutting teeth similar to those of modern piranha, but with a surprising evolutionary twist. Unlike any other known ray-finned fish, Serrasalmimus‘ teeth were true shearing carnassials anchored into bony sockets, with new replacement teeth forming directly below each current tooth – a very specific arrangement of features previously only known in mammals.
This is an especially remarkable example of convergent evolution because on land placental carnivorans were developing their own carnassials at the same time, just a few million years after the K-Pg mass extinction. Both mammals and pycnodonts were simultaneously taking advantage of the vacant predatory roles in their respective ecosystems, and ended up with incredibly similar tooth adaptations as a result.
First discovered in Jurassic-aged fossil deposits in Europe in the 1830s, this genus was quickly turned into a notorious wastebasket taxon for any similar-looking fossil fish. Over time dozens of different Lepidotes species were named, many of them rather dubious, from locations all around the world and spanning a time period of over 100 million years.
Lepidotes gigas
But despite Lepidotes being a wastebasket for almost two centuries, it wasn’t until surprisingly recently that any real progress began to be made on cleaning it all up.
In the early 2010s a large-scale review of ginglymodian relationships found that many “Lepidotes” species were either invalid or polyphyletic, belonging in completely different genera or families. True Lepidotes were restricted down to just the original type speciesLepidotes gigas and a few of its closest relatives, all from the early Jurassic of Europe, while some other forms were moved into the newer genera Scheenstia and Callipurbeckia. Since then some other “Lepidotes” have also been reclassified, creating new names like Macrosemimimus, Occitanichthys, and Quasimodichthys.
There’s still work needing to be done on untangling all these Lepidotes-like fish – Scheenstia might actually now represent several different lineages, for example – but at least Lepidotes itself is now in a much better situation than it was just a couple of decades ago.
Ornithoprion hertwigi here was one of the first eugeneodonts found with fossilized skull material, and helped with the early understanding of just how their weird jaw anatomy actually worked.
It lived during the Late Carboniferous, about 315-307 million years ago, in a shallow tropical sea that covered what is now southwestern Indiana, USA.
At only around 50cm long (~1’8″) it was one of the smaller eugeneodonts, and along with a small Helicoprion-like tooth whorl it also had a distinctive highly elongated chin. Similar to modern halfbeak fish this structure may have served a sensory function, helping Ornithoprion to detect prey in dark or murky waters.











{kind=link}