The distinctive pinhole eyes, leathery hood, and numerous tentacles of modern nautiluses were traditionally thought to represent the “primitive” ancestral state of early shelled cephalopods – but genetic studies have found that that nautiluses actually secondarily lost the genes for building lensed eyes, and their embryological development shows the initial formation of ten arm buds (similar to those of coeloids) with their hood appearing to be created via fusing some of the many tentacles that form later.
There’s a Cretaceous nautilidan fossil that preserves soft tissue impressions of what appear to be pinhole eyes and possibly a remnant of a hood, so we know these modern-style nautilus features were well-established by the late Mesozoic. But for much more ancient Paleozoic members of the lineage… we can potentially get more speculative.
So, here’s an example reconstructed with un-nautilus-like soft parts.
Up to about 20cm in diameter, (~8″), its shell featured long sideways spines which may have served as a defense against predators – or possibly as a display feature since they only developed upon reaching maturity.
Around 5cm long (2″), it had a downwards-curving egg-shaped shell that preserved the original color pattern on one fossil specimen, showing closely-packed crisscrossing vertical and horizontal bands.
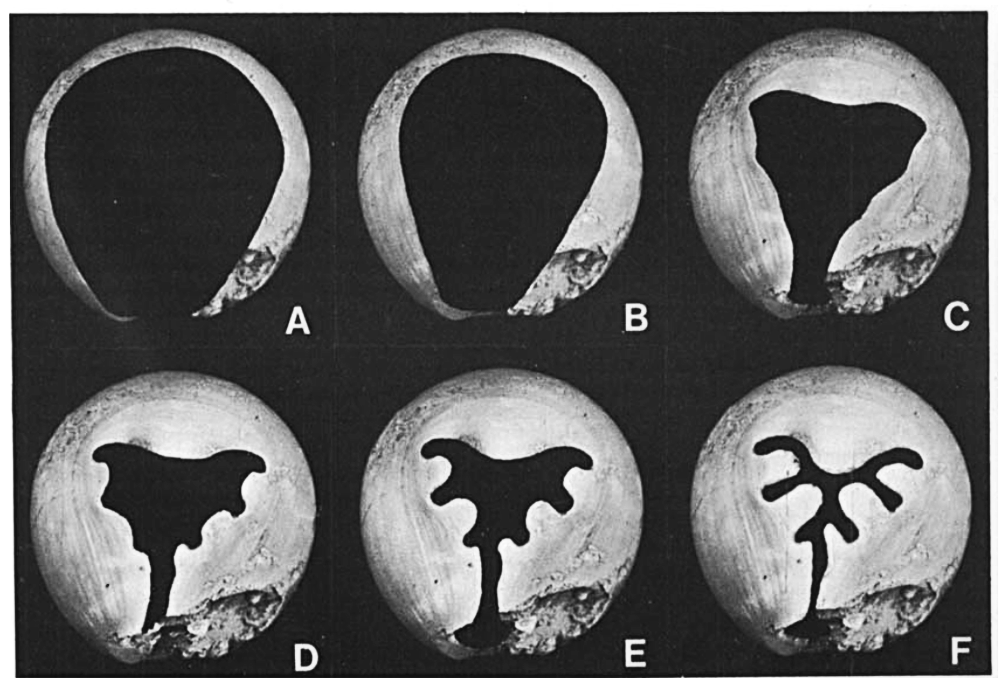
Like several of its close oncocerid and discosorid relatives, its shell also developed a highly constricted opening as it reached maturity. This eventually formed into a narrow visor-like shape with several lobes that probably correlated to the life positions of the eyes and arms, with a “spout” at the bottom for the siphon.
Development of the “visor” in the related Octameroceras sinuosum From fig 6 in Stridsberg (1981)
The function of this structure is still unclear. It may have been a defensive measure against predators – but it would have also severely limited the range of motion of the arms and the size of food that could be eaten through the mouth, suggesting that Hexameroceras may have specialized in very small prey, perhaps even filter-feeding.
Another possibility is that these visored nautiloids might represent brooding females, walling themselves into their shells to protect their eggs and dying after releasing the hatchlings through the tiny remaining gap.
Although the only nautiloids living today have characteristic tightly coiled shells, earlier in their evolutionary history these cephalopods were much more diverse.
And Glossoceras gracile here is an example of one of the more unusual groups of nautloids: the ascocerids.
Living during the Late Silurian, about 422 million years ago, in wheat is now Gotland, Sweden, Glossoceras was only around 5cm long as an adult (~2″). Like other ascocerids it started out its life looking like a fairly standard early nautiloid, with a long straight shell that curved slightly upwards, but as it approached maturity things got weird – the front part of the shell grew out into a much more bulbous flask-like shape, and the old juvenile section broke off entirely.
The gas-filled buoyancy chambers of its adult shell were positioned directly above its body chamber rather than behind like in other nautiloids, giving it very good stability in the water. The shell walls were also very thin and lightweight, which would have made it a much more maneuverable swimmer.
But one of the most bizarre of all was the genus Nipponites, whose ribbed shell looked like a bundle of tangled asymmetrical coils.
Nipponites bacchus lived in what is now Hokkaido, Japan, during the late Cretaceous about 90 million years ago. Around 10cm long (~4″), its shell was less tightly coiled up than its better-known relative Nipponites mirabilis, but these looser whorls were formed in the same way via a series of U-bends in different directions during its growth.
Despite their irregular and ungainly appearance, the unique shape of these ammonites seems to have actually been very hydrodynamically stable. They weren’t fast-moving, but they didn’t need to be, probably spending most of their time floating suspended in the water column catching small planktonic prey from around themselves.
But some bivalves break that arrangement, developing asymmetrical valves that can be massively different in size and shape.
Gryphaea arcuata was an oyster that lived during the Early Jurassic, about 200-174 million years ago, in the warm shallow seas that covered what is now Europe and eastern Greenland. Around 6cm long (~2.4″), its left valve was thick and strongly convex and curled, while the right valve was relatively thin and slightly concave forming a “lid”.
The gnarled curled claw-like shape of Gryphaea fossils led to them being colloquially known as “devil’s toenails” in some of the regions where they’re commonly found, with folk beliefs that they had the power to prevent joint pain.
Their shape was actually an adaptation to living on very soft seafloor sediments. The larger curled valve acted sort of like a boat on the soupy mud, supporting the Gryphaea‘s weight and preventing it from sinking.
The spiral-coiled shells of snails are their most familiar feature, giving them obvious external asymmetry, but gastropods are also defined by a specific type of internal asymmetry known as torsion.
Torsion is an anatomical process that occurs during larval development, and involves rotating their internal organs, mantle, and shell a full 180° relative to their head and muscular foot. This twists their gut into a U-shape, knots up their nervous system, and brings their respiratory organs and anus up close to their head.
And we still don’t really know why they do it.
One idea (the “rotation hypothesis”) is that it originated as a defensive function after early gastropods began developing their spiral shells. The shell opening may have originally been positioned at early gastropods’ rears, meaning they retracted their bodies back-end-first leaving their heads and sensory structures still vulnerable – but twisting the shell around would allow them to pull their front end in faster instead.
A competing idea (the “asymmetry hypothesis“) instead proposes that the shape of the coiled shell restricted the gills of early gastropods, which may have originally been positioned in mantle cavities on each side of their bodies. In response to this they developed a single larger gill cavity on just one side of their body, and then gradually expanded and rotated this asymmetric feature around to the front for better aeration.
In either case this resulted in some of the rest of their anatomy “coming along for the ride”. And regardless of whatever the original evolutionary advantage of torsion actually was, it made gastropods incredibly successful – they’re a massively diverse group, second only to the insects in terms of sheer number of species, and today they’re found all over the world in almost every habitat from deep sea trenches to high mountain elevations.
Spinyplatyceras arkonense
Spinyplatyceras arkonense lived in what is now Ontario during the mid-Devonian, about 391-385 million years ago. Around 5cm long (2″), it was part of a group of Paleozoic marine snails known as platyceratids, which were probably related to either modern limpets or neritomorphs.
Platyceratids seem to have had a unique parasitic relationship with crinoids, attaching themselves to the top of the host’s body and using their radula to drill into them, either robbing food directly from the crinoid’s gut or feeding on its other internal organs.
The long spines on Spinyplatyceras‘ shell probably helped to deter predators. In an interesting case of coevolution the crinoid hosts of some platyceratids developed their own defensive spines, too – and it seems this wasn’t to prevent the snails from infesting them, but to also discourage the snails’ predators. These crinoids may have been frequently indirectly injured during snail-eating predators’ attacks, and it might have actually “cost” them less to keep enduring an infestation than to deal with the collateral damage of the snails being removed.
First discovered in the early 1970s, in the mid-CarboniferousBear Gulch Limestone deposits (~324 million years ago) of Montana, USA, it was initially mistaken for the long-sought-after “conodont animal” due to the presence of numerous conodont teeth inside its body. But just a few years later well-preserved eel-like conodont animals were found elsewhere, and it became apparent that the conodont teeth inside Typhloesus had actually just been part of its last meal.
But if it wasn’t a conodont… then what was it?
Up to about 10cm long (4″), Typhloesus had a streamlined body with a vertical tail fin and paired “keels” along its sides. It had a mouth and a gut cavity, but no apparent anus, and it also didn’t seem to have any eyes or other sensory structures. And in the middle of its body there was something very weird – a pair of “ferrodiscus” organs, disc-shaped structures which contained high concentrations of iron but whose function was completely unknown.
This anatomy just didn’t match any other known animals, so much so that it gained the nickname of “alien goldfish”.
For the next few decades it remained a bizarre enigma, at best tentatively considered to represent an unknown lineage of some sort of metazoan that left almost no other fossil record due to being entirely soft-bodied.
But now, 50 years after its initial discovery, we might just finally have a clue about Typhloesus’ true identity.
It’s not a definite identification yet, and even if it was a mollusc it was an incredibly strange one, with features like the ferrodiscus still lacking any explanation. But this discovery at least shows that there are still new details waiting to be found in the “alien goldfish” fossils, and gives us a start towards bringing its classification back down to earth.
Heteromorphs’ strange shells would have created a lot of drag in the water, and they may not have been especially agile swimmers, but they were very hydrodynamically stable and easily maintained neutral buoyancy. Their paleobiology has only just started to be properly understood in recent years, and now most species of these ammonites are thought to have floated suspended in the photic zone and twilight zone of the open ocean, catching small zooplankton from the water around themselves.
What these ammonites were doing obviously worked very well for them, because they were incredibly diverse and successful during the Cretaceous period. They were also the only type of ammonite to persist for a short time after the end-Cretaceous mass extinction, existing as a “dead clade walking” for another half a million years or so before finally disappearing entirely.
Hamites attenuatus
The hamitids were a group of heteromorphs from the mid-Cretaceous (~110-90 million years ago), with their namesake genus Hamites traditionally being used as a wastebasket taxon for anything that that didn’t neatly fit into any other group of similar heteromorph ammonites.
By the late 1990s Hamites had become a mess of multiple different diverse lineages, with over 20 species all lumped together – and this was a problem because the hamitids were the ancestors of severalotherheteromorphammonitelineages, and having the taxon in such disarray made studying the evolutionary origins of all those other groups very difficult.
So in the early 2000s attempts were made to clean this all up, figuring out the relationships between the different Hamites species and dividing the genus into multiple new genera.
There hasn’t been much more detailed research on the relationships of hamitids since then – and other groups of heteromorphs are still in need of revision – but it’s a start at clearing the wastebasket, at least.
Bellerophonts were small snail-like marine molluscs that were either early gastropods or very close relatives of them. They had symmetrically-coiled shells superficially shaped like those of nautiluses, with about half of the shell covered by their mantle similarly to some modern seasnails, and some fossil shells also preserve hints of banded color patterns.
First appearing in the late Cambrian (~500 million years ago), these molluscs existed all the way until the early Triassic, surviving the Great Dying mass extinction (~252 million years ago) only to go extinct just a short time later (~249 million years ago) – a phenomenon known as “dead clade walking”, when a group just barely scrapes through a mass extinction event but doesn’t manage to actually recover afterwards.
The whole group is something of a wastebasket of similar-looking shells, and might actually be more of an “evolutionary grade” made up of various early gastropods and gastropod-relatives than a single defined lineage.
But there’s also another wastebasket nestled inside this wastebasket: the namesake of them all, the genus Bellerophon.
Bellerophon tenuifascia
Originally named in 1808, this genus has had a huge number of species assigned to it over the last couple of centuries. This gives a false impression that Bellerophon-like molluscs didn’t change for hundreds of millions of years, and it makes figuring out their actual long-term patterns of evolution and extinction much more difficult.
In the last few decades some mollusc paleontologists have been gradually chipping away at Bellerophon, and multiple new genera have been broken off from it. But even today it remains a very bloated mess – there are still well over a hundred named species spanning about 230 million years of geologic time.
Studies do indicate the whole genus is highly polyphyletic, made up of a tangle of multiple different lineages that all really need to be revised and renamed – but there’s a lot of work still needing to be done to clean up this particular wastebasket.















{kind=link}
{kind=link}
{kind=link}