

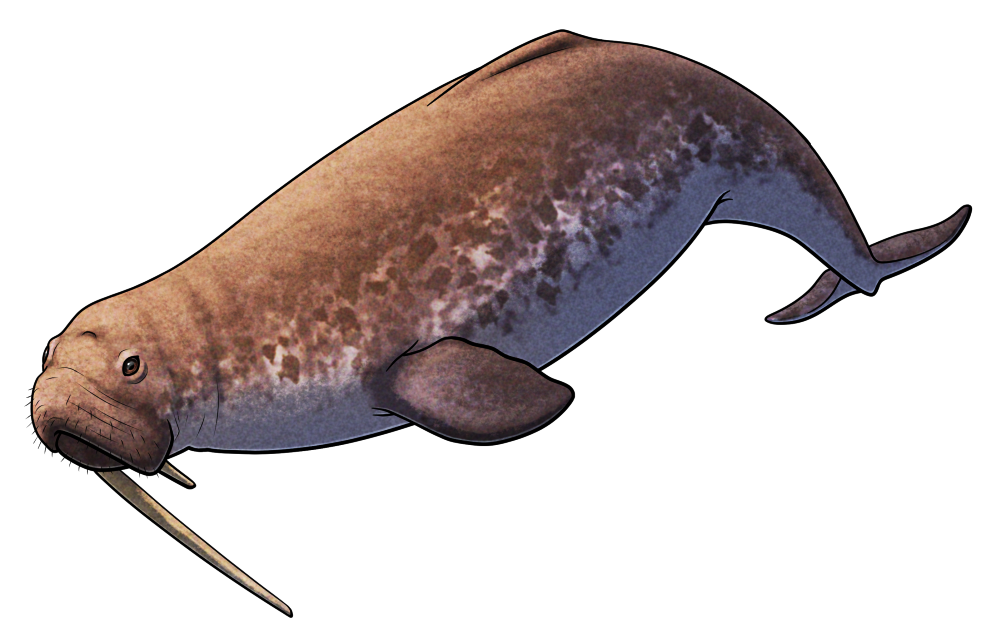
Albireonids were an early branch of the delphinoid whales, with their closest living relatives being modern oceanic dolphins, narwhals and belugas, and porpoises. Known from temperate latitudes of the North Pacific Ocean between the late Miocene and the late Pliocene, about 9-2.5 million years ago, their fossil remains are very rare in coastal deposits and they seem to have primarily been offshore open ocean animals.
Albireo whistleri is the best known member of this family, represented by a near-complete skeleton from what is now Isla de Cedros in Baja California, Mexico, dating to the late Miocene between about 8 and 6 million years ago. It was a rather small dolphin, around 2.5m long (~8’2″), with a stocky body, fairly broad flippers, and skull anatomy with some convergent similarities with the modern Dall’s porpoise.
Interestingly these dolphins also seem to have frequently had pathological neck vertebrae, with both Albireo whisteri and the younger species Albireo savagei from California, USA, showing unusually asymmetrical atlas bones – but on opposite sides to each other. This might be due to illness or injury earlier in life, or possibly be evidence of some sort of “handedness” with individuals preferring to perform some actions more with one side of their body than the other.
Continue reading “Albireo”









{kind=link}
{kind=link}
{kind=link}