

The dodo wasn’t the only unique bird to evolve on Mauritius. While about eight other endemic bird species still survive today, there were at least twice that many before the arrival of humans in the late 1500s – including the broad-billed parrot Lophopsittacus mauritianus.
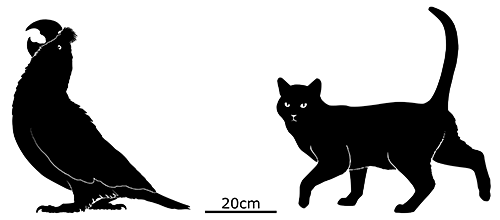
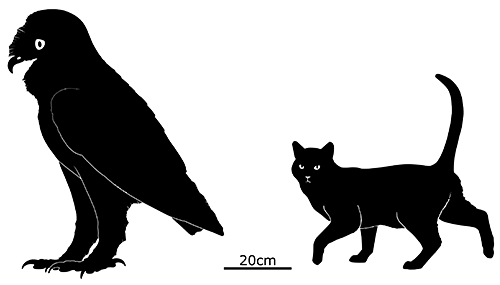
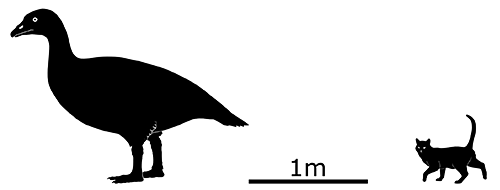
Also referred to as the “Indian raven” in historical accounts, it was a fairly large bird measuring between 45 and 65cm in length (1′6″-2′1″). Unusually for a parrot it had a high degree of sexual dimorphism, with males being significantly bigger than females.
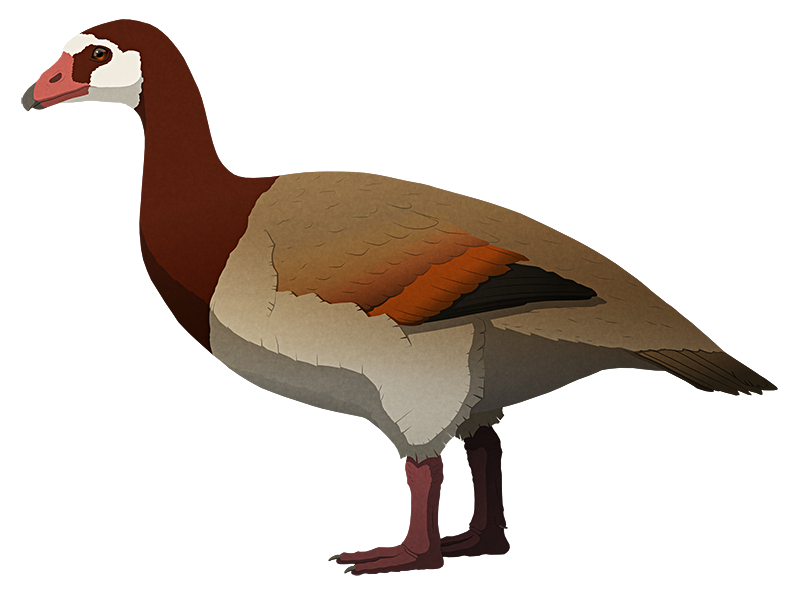
Many images depict it as entirely black or blue-grey, but this seems to be based on a misinterpretation. More recent translations of old Dutch descriptions suggest it was actually much more colorful, with a red beak, blue head, and reddish body.
It had a proportionally big head and a flattened skull, and seems to have had a highly specialized diet, using its its large strong beak to crack open hard seeds and nuts like modern hyacinth macaws.
It was near-flightless, capable of taking to the air only with difficulty, and was said to be “bad-tempered”. Attempts to keep individuals in captivity failed, the birds refusing to eat, and while the wild population had apparently learned to be wary of humans by the late 1660s by that point it was already too late. Much like the dodos they lived alongside, a combination of deforestation, hunting, and predation by invasive mammal species sent them into extinction by the 1680s.












.jpg){kind=link}
{kind=link}