Although it was a distant evolutionary cousin to plesiosaurs (and even more distantly to modern turtles), it was actually most closely related to an early sauropterygian lineage known as the pachypleurosaurs – a group of small lizard-like aquatic reptiles with tiny heads, long necks, and paddle-like limbs.
It had an unusually short and rounded-off snout compared to its relatives, and since it would have lived alongsidemanyotherdiversemarinereptiles it was probably specialized for a slightly different ecological niche.
Nanodobenus arandai, a pinniped from the mid-to late Miocene (~16-9 mya) of Baja California Sur, Mexico. Although it would have looked very similar to a sea lion, it was actually an early member of the walrus lineage that lacked the specialized long tusks that characterize its modern relatives.
At just 1.65m long (5′5″) it was only about half the size of living walruses, making it the smallest member of the group ever discovered and leading to it being given the nickname “smallrus”.
It probably occupied a similar sort of fish-eating ecological niche as true sea lions – which eventually replaced it in the region after its extinction – and since it lived alongside several other larger species of walrus it may have become dwarfed to avoid direct competition with them.
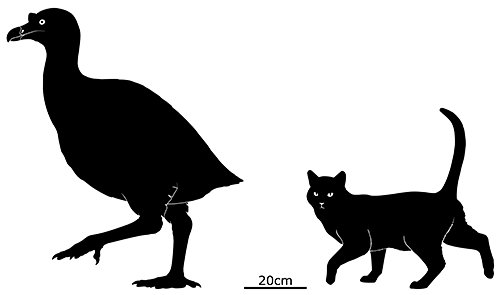
Standing around 50cm tall (1′8″), it had a slender body, long legs, a long neck, and a narrow goose-like beak. It also had an unusual pair of bony bumps on its skull which may have supported some sort of small crest superficially similar to the knob on the head of the modern magpie goose.
Temperatures in Antarctica at the time were much warmer than today, and the area where its fossils were found would have been a temperate estuary or river delta. It was probably an omnivorous wading bird, feeding on vegetation, small fish, and invertebrates in shallow freshwater.
Although it somewhat resembled a presbyornithid it was actually part of an even earlier branch of the waterfowl evolutionary tree – so its ancestors must have originated much further back in the Late Cretaceous – and their similar body shapes hint that the common ancestor of all waterfowl may also have been a rather leggy bird. Conflicto’s closest known relative might actually be the similarly-aged Anatalavis (which was previously though to be a primitive magpie-goose) from North America and Europe, suggesting that its lineage was quite widespread and already taking advantage of vacant niches in the immediate wake of the Cretaceous-Paleogene mass extinction.
Much like Japan, ancient Flores had a succession of dwarf stegodontids – close relatives of modern elephants that were capable of island-hopping through Indonesia by swimming.
Stegodon sondaari
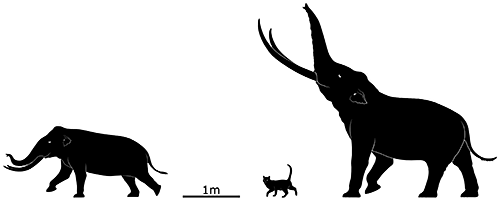
Stegodon sondaari lived on Flores during the Early Pleistocene, about 900,000 years ago, and was the size of a small water buffalo at just 1.2m (3′11″) tall at the shoulder. It was probably descended from the larger Stegodon trigonocephalus, known from Java, and it had proportionally short legs which may have been an adaptation to clambering over rough terrain and steep inclines.
Around 850,000 years ago Stegodon sondaari disappeared from Flores, probably due to a large volcanic eruption, but a new wave of stegodontids quickly recolonized the island. The mid-sized Stegodon florensis probably originated from either Java to the west or Sulawesi to the north, and eventually evolved into a new dwarfed subspecies.
Stegodon florensis insularis
Stegodon florensis insularis wasn’t quite as small as its predecessor, standing around 1.8m tall (5′10″). It probably didn’t shrink quite so much due to the existing presence of various predators on Flores, since it was likely the main prey of large Komodo dragons, it was hunted by Homo floresiensis, and it may also have been occasionally targeted by giant storks.
It seems to have disappeared around the same time as several other unique endemic species, between 50,000 and 20,000 years ago, due to either climate change, another volcanic eruption, or the arrival of modern humans – or perhaps a combination of all of those factors.
And that’s all for this month… but Island Weirdness will be back later for part 2, with more giants, more dwarfs, and so many elephants.
Standing at an adult height of about 1.1m tall (3′7″), they were smaller than any population of modern humans and are thought to represent an unusual case of insular dwarfism.
They also had much smaller brains than would be expected for their size, similarly to the miniature hippos of Madagascar, which was probably an energy-saving adaptation in an island environment with limited resources, since brains are metabolically expensive organs. An area of their brains associated with higher cognition was about the same size as in modern humans, however, so they weren’t necessarily less intelligent – stone tools and butchery marks on dwarf elephant bones suggest they were cooperatively hunting, and there’s also possible evidence of fire use for cooking.
It’s not clear exactly where they belong on the human family tree, and attempts at extracting DNA from the known remains have so far failed. They might be descendants of a population of Homo erectus who arrived on Flores about 1 million years ago, or they may even have been part of a much older unknown lineage that dispersed from Africa over 2 million years ago.
Although they were initially thought to have lived on Flores from 190,000 years ago up to about 12,000 years ago, more accurate dating of the cave where their skeletal remains were discovered suggests they actually disappeared about 50,000 years ago – about the same time that modern humans arrived on the island.
Indonesia is located at the junction between several tectonic plates, and as a result a large number of volcanic islands make up the region. While some of these islands have had land connections to Asia or Australia in the past, others are separated by deep ocean trenches and have been isolated with little movement of species between them.
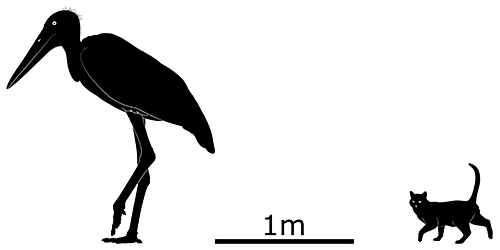
Leptoptilos robustus was a huge stork, closely related to the living marabou and adjutants but estimated to have been at least 20% larger. It would have stood around 1.8m tall (5′10″) and had a chunkier build with unusually heavy thick-walled bones, suggesting it may have become functionally flightless. Only fragmentary arm bones were found, however, so its unknown whether its wings were reduced in size or not.
There were few large carnivorous mammals on Flores (possibly none), and Leptoptilos robustus would have had little competition for carrion and prey. It may even have filled an ecological niche similar to the giant Hatzegopteryx of Cretaceous Hațeg Island – a large terrestrial stalking predator eating any smaller animals unfortunate enough to fit into its mouth.
The known remains date to the Late Pleistocene, around 50,000 to 20,000 years ago, and this giant bird seems to have gone extinct sometime during that date range. It’s unclear what killed it off, but possible factors include a changing climate on the island, a major volcanic eruption, and the arrival of modern humans.
Located to the east of Mauritius, the small island of Rodrigues is the geologically youngest of the Mascarenes, formed only about 1.5-2 million years ago.
And it also had its own large flightless bird – the Rodrigues solitaire, Pezophaps solitaria.
It was closely related to the dodo, although it wasn’t a direct descendant. Based on DNA studies their last common ancestor is estimated to have lived about 20 million years ago, so they must have each convergently evolved from separate pigeon lineages that arrived on each island at different times.
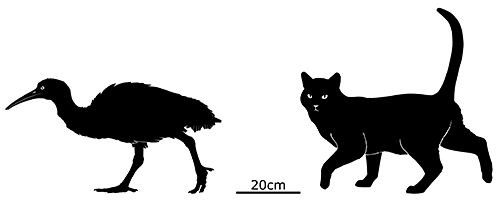
Standing 70-90cm tall (2′4″-2′11″), with males being larger than females, it had long legs, a long neck, and a slightly hooked beak with a black band described as resembling a widow’s peak. Its plumage was grey and brown, and it was reportedly aggressively territorial and capable of giving a strong bite.
It had large lumpy bony knobs on its wrist bones that were used as weapons to clobber each other while fighting. Due to this its wings were less reduced than the dodo, retaining stronger musculature, and it was apparently also capable of using them to create loud low-frequency sounds for communication – possibly in a similar manner to modern crested pigeons’ whistling wings.
The solitaire survived for longer than both its dodo cousin and the Réunion ibis, but only because its island was rarely visited by humans until the late 1600s. Once Rodrigues began to be exploited the story became the same as the other Mascarene islands: a combination of hunting, habitat destruction, and predation by invasive species rapidly dwindled its population. It likely went extinct sometime between the 1730s and 1750s, since an exhaustive search for a live specimen in 1755 failed to find a single bird.
The island of Réunion was formed southwest of Mauritius ony about 2-3 million years ago, and likewise developed its own vertebrate ecosystem from various birds, reptiles, and bats arriving via island hopping and rafting.
Accounts from the 1600s and 1700s of a white bird on the island known as the “Réunion solitaire” were for a long time thought to refer to a close relative of the dodo, but no dodo-like subfossil remains were ever found.
Eventually bones of a different type of bird were discovered: an ibis. Closely related to the African sacred ibis, Threskiornis solitarius was a similar size at about 65cm long (2′2″) but had a much chunkier build. Its beak was shorter and straighter than other ibises, and it had reduced flight capabilities – features that matched the old solitaire descriptions surprisingly well.
Its coloration was mostly white, merging into grey and yellow, and it had glossy or iridescent black ostrich-like plumes on its rear. It mainly lived in forests and used its beak to search through soft soil for invertebrates like worms and insects.
A combination of its island tameness and the fact that it was considered to be good eating resulted in it being heavily hunted during the 1600s, and along with pressure from invasive mammals – such as predation by cats and its eggs being eaten by pigs – it soon became a rare sight, found only in more remote highland areas of the island.
The last definite account of the Réunion ibis was in the early 1700s, and it was probably completely extinct within the first couple of decades of that century.
Along with its uniquebirds, Mauritius was also home to many endemic reptile species. In the absence of terrestrial mammals giant tortoises were the largest herbivores on the island, and various geckos, skinks, and snakes helped to fill out the rest of the vertebrate ecosystem.
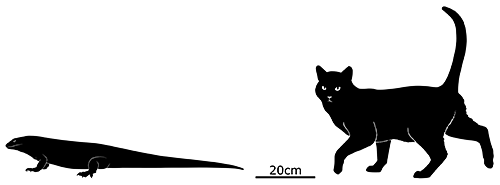
Leiolopisma mauritiana was a very largeskink, one of the biggest ever known to have existed with a total length of around 80cm (2′7″). Its ancestors originated in Australasia, over 5600km away (~3500 miles) at least 3-4 million years ago – and they must have endured a particularly long ocean rafting journey without any island hopping stops, since none of the other islands along that route seem to have ever had populations of similar skinks.
It probably lived in rocky areas, possibly also being capable of digging burrows, and would have eaten an omnivorous diet of seeds, fruits, invertebrates, and smaller lizards and birds.
By the early 1600s it was already extinct, very soon after the arrival of humans, probably due to predation from invasive mammals like rats. However, its half-sized close relative Leiolopisma telfairii does still survive on rat-free Round Island a short distance to the north of Mauritius, and recent conservation efforts have been rebuilding its population and setting up new colonies on other nearby small islands.



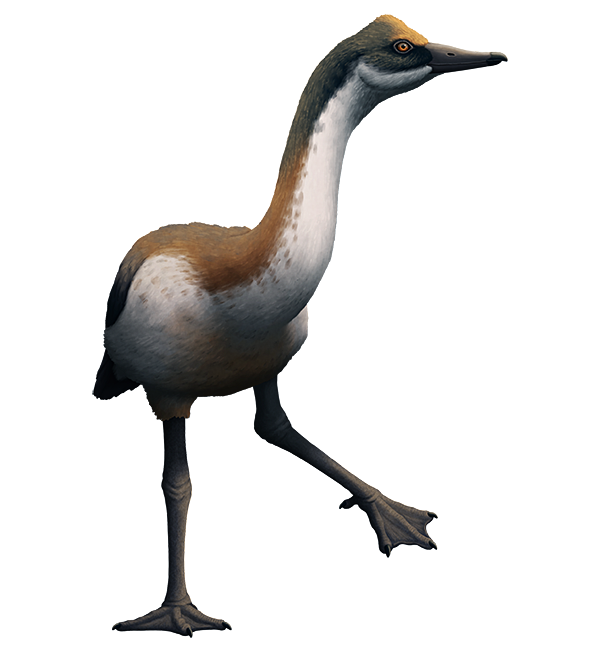









{kind=link}