The first definite ichthyosaur fossil found in Colombia was a single well-preserved skull, found in Early Cretaceous deposits dating to between 130 and 112 million years ago.
Although first discovered in the 1970s, this marine reptile wasn’t described until the late 1990s, at the time being named as a species of Platypterygius. But since then more pieces of the skeleton have been recovered, and the Platypterygius genus has been found to be a wastebasket taxon in need of revision, so in 2021 the Colombian ichthyosaur got a more detailed redescription and its own distinct name: Kyhytysuka sachicarum.
Kyhytysuka was a mid-sized ichthyosaur, about 5.5m long (18′) – about the size of a small modern orca – with a large head and a long robust snout. Its teeth varied in size, shape, and spacing along its jaws, with several different regions that were specialized to catch, slice, and crush its prey.
It could also open its jaws very widely, possibly up to an angle of 75°, suggesting it was able to tackle particularly large prey such as other marine reptiles. Possible soft tissue preservation around its lower jaw might also be evidence of elastic connective tissue that would have allowed its throat to expand out while swallowing big prey items.
This makes Kyhytysuka the first known example of a Cretaceous-aged ichthyosaur with an apex predator lifestyle, convergently evolving a similar ecological role to some earlier Triassic and Jurassic species.
First discovered in the early 1970s, in the mid-CarboniferousBear Gulch Limestone deposits (~324 million years ago) of Montana, USA, it was initially mistaken for the long-sought-after “conodont animal” due to the presence of numerous conodont teeth inside its body. But just a few years later well-preserved eel-like conodont animals were found elsewhere, and it became apparent that the conodont teeth inside Typhloesus had actually just been part of its last meal.
But if it wasn’t a conodont… then what was it?
Up to about 10cm long (4″), Typhloesus had a streamlined body with a vertical tail fin and paired “keels” along its sides. It had a mouth and a gut cavity, but no apparent anus, and it also didn’t seem to have any eyes or other sensory structures. And in the middle of its body there was something very weird – a pair of “ferrodiscus” organs, disc-shaped structures which contained high concentrations of iron but whose function was completely unknown.
This anatomy just didn’t match any other known animals, so much so that it gained the nickname of “alien goldfish”.
For the next few decades it remained a bizarre enigma, at best tentatively considered to represent an unknown lineage of some sort of metazoan that left almost no other fossil record due to being entirely soft-bodied.
But now, 50 years after its initial discovery, we might just finally have a clue about Typhloesus’ true identity.
It’s not a definite identification yet, and even if it was a mollusc it was an incredibly strange one, with features like the ferrodiscus still lacking any explanation. But this discovery at least shows that there are still new details waiting to be found in the “alien goldfish” fossils, and gives us a start towards bringing its classification back down to earth.
A wastebasket taxon is what happens when species can’t be easily classified and instead get hurled into a “catch-all” category.
…But that’s not the only kind of taxonomic tangle that can befall a new discovery.
When a scientific name is assigned to a new species, but it isn’t given a corresponding formal description and type specimen, it becomes a nomen nudum – a “naked name”. Without a proper description and assigned holotype the name isn’t valid, and the new species isn’t technically accepted by the wider scientific community.
This has even happened to some surprisingly famous names. In the 1920s Velociraptor mongoliensis was briefly given the nomen nudum“Ovoraptor djadochtari” before getting its much more familiar name when it was officially described. Meanwhile the giant pterosaur Quetzalcoatlus northropi was stuck as a nomen nudum for decades, only finally getting a proper published description in 2021.
And there’s another particular long-standing nomen nudum that became mildly infamous – “Thotobolosaurus”, the “trash heap lizard”.
Kholumolumo ellenbergerorum
Discovered next to a literal trash pile in the village of Maphutseng in Lesotho, a few scattered and broken bones of this “prosauropod” sauropodomorph dinosaur were first found in 1930. But it wasn’t until the mid-1950s that a more extensive bonebed began to be unearthed at the site, and over the next decade over 1000 fossil fragments were collected.
In the mid-1960s the remains were initially classified as belonging to Euskelosaurus browni (which is now considered to be a wastebasket taxon), but just a few years later in 1970 the “Maphutseng Beast” was re-evaluated as a species new to science. It was referred to as “Thotobolosaurus mabeatae” – based on the local name of the discovery site, “Thotobolo ea ‘Ma-Beata” (trash heap of Beata’s mother) – but this name was never actually formally published.
Despite “Thotobolosaurus” being an undescribed nomen nudum it nonetheless went on to be repeatedly referenced in scientific literature over the next few decades, and appeared in several popular dinosaur books (even as recently as 2020!).
In the mid-1990s it was alternatively named “Kholumolumosaurus ellenbergerorum” in a Ph.D. dissertation, with this name derived from the kholumolumo, a reptilian creature in Sotho mythology, and the Ellenberger brothers who worked on the site. But this also didn’t count as a formal publication and instead became a second nomen nudum for the species.
Eventually, 90 years after the first bones were found and 50 years after the debut of the name “Thotobolosaurus”, this long-neglected sauropodomorph was finally given a proper published full anatomical description in 2020.
And it also got a third name, this time officially valid, based on the second one from the 1990s: Kholumolumo ellenbergerorum.
For something associated with trash for so long, Kholumolumo is actually now one of the most completely-known prosauropods. At least five different individuals were present in the collected fossil material, possibly as many as ten, and between them most of the full skeleton is represented – with the exception of the skulls, which are only known from a couple of small fragments.
We now know Kholumolumo was rather heavily-built, with chunky limb bones and unusually short shinbones. It would have been one of the biggest animals around in the Late Triassic (~210 million years ago), measuring at least 9m long (~30′) and weighing around 1.7 tonnes (1.9 US tons), but despite its size it seems to have still been bipedal.
Due to the highly disarticulated nature of the bones the fossil site may have been a “bone accumulation area”, a place where dismembered bits and pieces of different carcasses were regularly carried to be eaten by a predator or scavenger – essentially a trash heap, fittingly enough. A couple of “rauisuchian” teeth have actually been found among the remains, which might indicate what was chomping on these particular Kholumolumo.
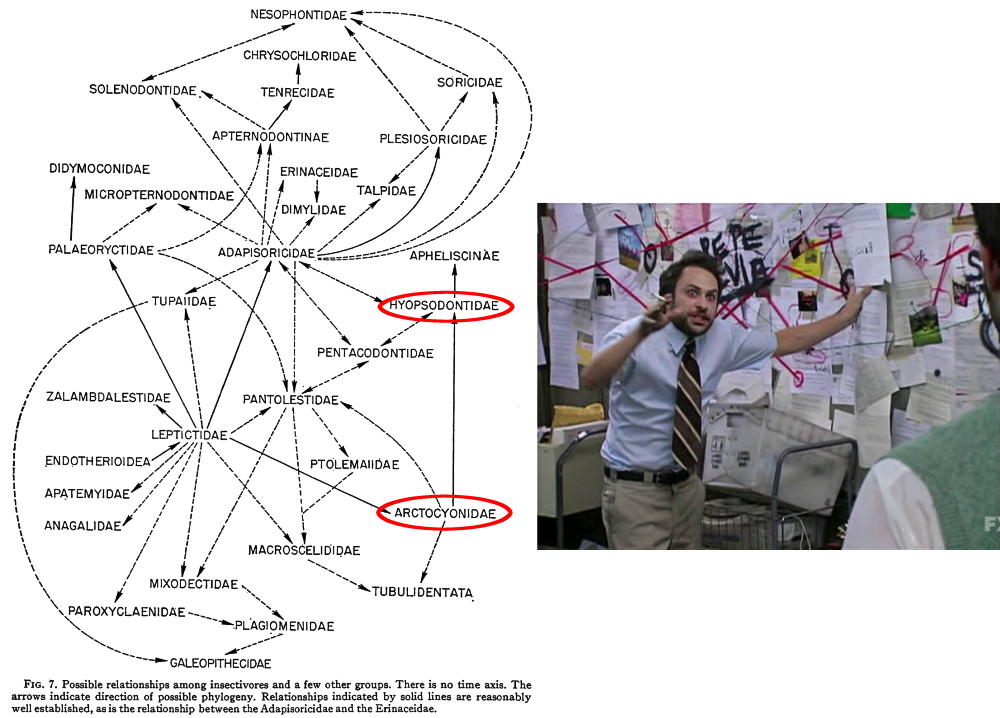
“Insectivora” was a wastebasket taxon so bad it had to be revised multiple times, but there’s another particularly infamous case in mammal taxonomy that’s still in the process of being resolved – the “condylarths“.
This group was first created in the early 1880s, during the Bone Wars, and initially was just a subgroup of odd-toed ungulates containing only the phenacodontids. But just a few years later Condylarthra was promoted up to its own order, and groups like the periptychids and hyopsodontids were added in too.
Then over the next few decades various groups were added and removed from the condylarths, most notably with the mesonychids and arctocyonids being brought in from their previous position with the creodonts.
By the mid-20th century the condylarths had become a big convenient dumping ground for any and all “primitive” ungulate-like mammals that didn’t easily fit into any modern groups, ranging in age from the early Paleocene through to the early Oligocene. But it soon became apparent that they had the same problem as the “insectivores” – there weren’t really any unique anatomical features that united all these animals together.
They generally had rounded-cusped molar teeth and hoof-like toes, but they also had rather generalized “primitive mammal” features and a diverse range of ecologies. Some were small herbivores, but others were coati-like or dog-like omnivores, and some were even bear-sized carnivores.
From left to right, top row: Hyopsodus lepidus (hyopsodontid), Meniscotherium chamense (phenacodontid), Arctocyon primaevus (“arctocyonid”). Bottom row: Ectoconus ditrigonus (periptychid), Mesonyx obtusidens (mesonychid)
It wasn’t even clear how the various different condylarth groups were actually related to each other. The best guess was that arctocyonids had arisen from within the “insectivores”, with a Protungulatum-like form as the common ancestor of all the other condylarths. Where exactly modern ungulates had then evolved from within the condylarths was also still uncertain.
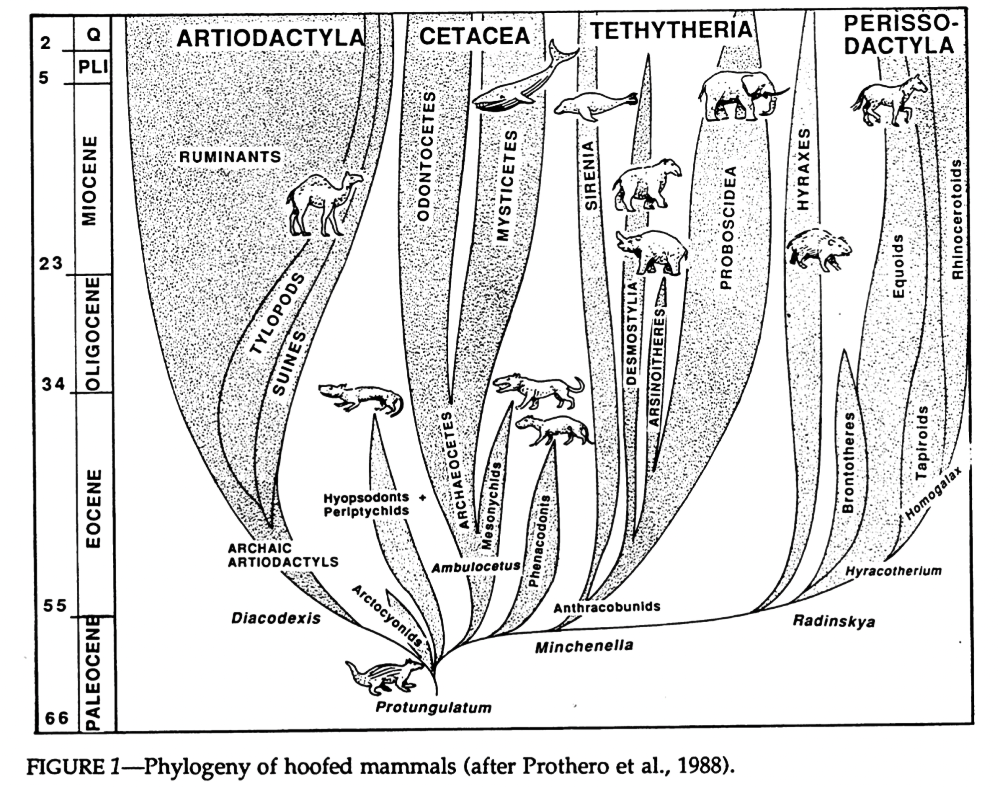
Cladistic analysis in the 1980s began to tackle the confusing pile of assorted condylarths, and showed that they weren’t the single ancestral source of all modern ungulates, but instead a loose collection of several unrelated groups from all over the ungulate evolutionary tree. Arctocyonids, periptychids, and hyopsodontids were placed as early “primitive” lineages, phenacodontids were loosely linked with the ancestors of odd-toed ungulates once again, and mesonychids were considered to be the ancestors of whales.
And, once again paralleling the mess of the “insectivores”, it wasn’t until genetic methods became available in the late 1990s that larger-scale ungulate relationships began to be properly resolved. The paenungulates (elephants, hyraxes, and sirenians), which had been traditionally considered to be a major branch of ungulates, were removed entirely and reclassified as afrotheres. And, along with some new fossil discoveries, whales were recognized as having actually evolved from within the even-toed ungulates instead of from mesonychids.
This shake-up threw the still-problematic “condylarth” classifications back into question – with some “condylarths” turning out toalso be afrotheres instead of true ungulates.
Today the actual relationships of the main “condylarth” ungulate families are still in the process of being figured out, and there’s a lot of remaining uncertainty and disagreement about them.
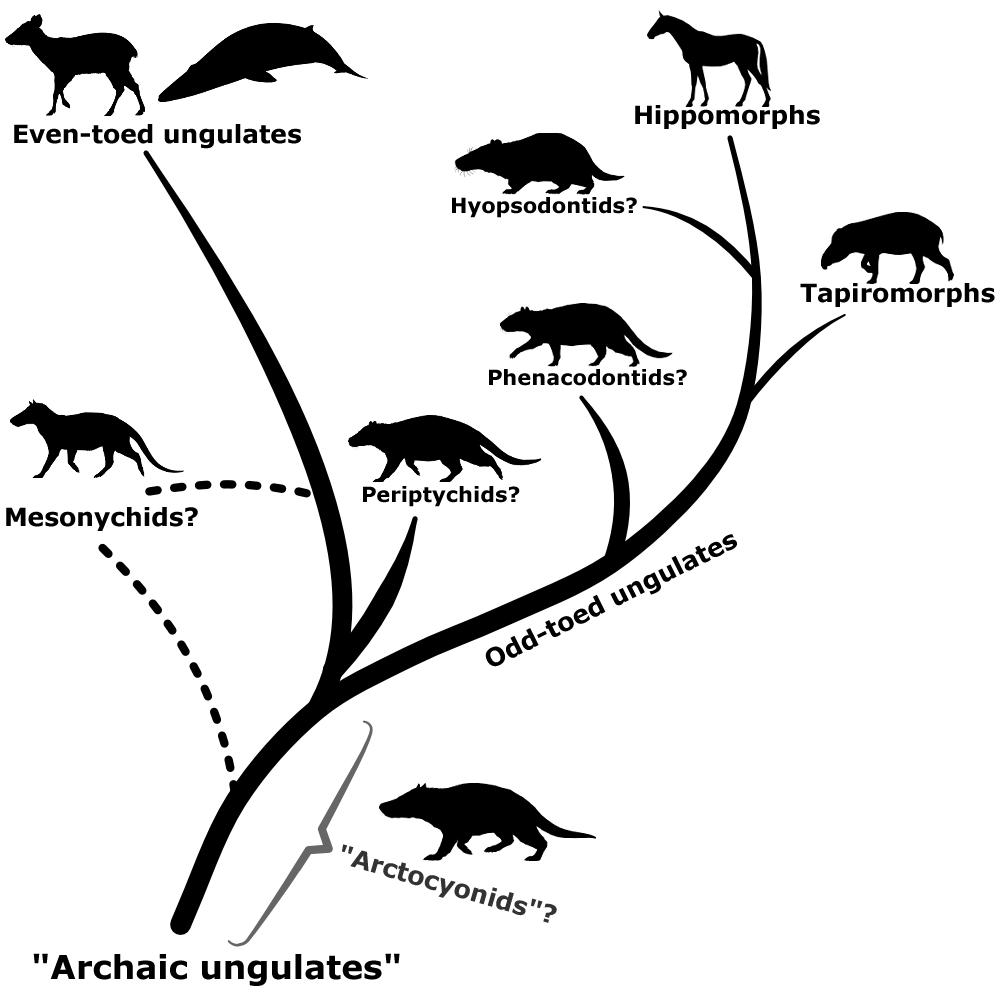
Phenacodontids seem to have mostly maintained their traditional position as early odd-toed ungulates, and hyopsodontids may potentially be part of this group too – possibly as members of the hippomorph lineage, closely related to horses and brontotheres. Arctocyonids might be a wastebasket themselves, with some studies finding them to be a mix of several different archaic ungulate lineages. Periptychids may have links to the even-toed ungulates. The mesonychids, meanwhile, are now generally considered to be a separate order from the traditional “condylarths”, and may be either an early branch of the even-toed ungulates or much more basal ungulates closely related to the “arctocyonids”.
Since the term “condylarth” no longer has any real taxonomic meaning some paleontologists have proposed replacing it with “archaic ungulate” to distance from the historical messiness of the old name. But this hasn’t really caught on, and many papers still use “condylarth” in a very loose sense to refer to an “evolutionary grade” of early ungulates of unclear evolutionary affinities.
And while that’s the last main entry for this month, we’re not quite done yet. There’s still one weekday left in October, and after digging through so many taxonomic garbage cans there’s only one place we can go now.
The Ptychopariida were some of the earliest known trilobites, first appearing in the early Cambrian about 521 million years ago and surviving until the end of the Ordovician about 444 million years ago. They included some of the most numerous and common trilobite species, and were probably ancestral to multiple other major lineages – including the very last trilobites at the end of the Permian – making them incredibly important in understanding the overall evolution of trilobites as a whole.
…But this group is also one of the biggest wastebaskets in paleontology.
First established in the early 20th century, the ptychopariids seemed to have some fairly good defining characteristics based on their facial sutures, large thoraxes, and relatively small pygidia. But the group quickly became a dumping ground for a massive amount of Cambrian trilobites, eventually containing numerous different families, hundreds of genera, and many more individual species.
Actually figuring out their internal evolutionary relationships also turned out to be extremely difficult – so much so that some paleontologists working on them just gave up trying and arranged the genera names alphabetically instead!
Even cladistic studies from the 1970s onward struggled to make sense of these highly “problematic” trilobites, and any larger-scale analysis was a daunting task due to how huge and diverse the ptychopariid wastebasket had become over the years. Worse, some of the anatomical features the group had been based around were starting to look more like the result of a lot of convergent evolution across multiple lineages than any actual shared ancestry.
Efforts were still made at breaking up the mess, however, with better-understood sub-groups like the Proetida, Harpida, Asaphida, Trinucleida, and Olenida being gradually split off into their own separate orders over the course of the last few decades.
Ptychoparia striata
But even by the early 2010s what remained of the Ptychopariida was still paraphyletic at best, more of an “evolutionary grade” of early trilobites than a single lineage, with most of its constituent families also rather poorly defined. There was even a proposal to abandon the group entirely, stating that “it serves no scientific purpose” and that its orphaned contents should be considered “order uncertain” until their actual relationships can be untangled.
Today the “ptychopariids” are in dire need of a full revision – since they were the ancestors of many other major groups they’re still crucial for understanding early trilobite evolution. There may be a salvageable single lineage somewhere in the remains of this wastebasket, even if it’s restricted to just close relatives of the genus Ptychoparia, but until somebody tackles them properly they’re stuck in taxonomic limbo with their name only being used in a loose sense.
First discovered in Jurassic-aged fossil deposits in Europe in the 1830s, this genus was quickly turned into a notorious wastebasket taxon for any similar-looking fossil fish. Over time dozens of different Lepidotes species were named, many of them rather dubious, from locations all around the world and spanning a time period of over 100 million years.
Lepidotes gigas
But despite Lepidotes being a wastebasket for almost two centuries, it wasn’t until surprisingly recently that any real progress began to be made on cleaning it all up.
In the early 2010s a large-scale review of ginglymodian relationships found that many “Lepidotes” species were either invalid or polyphyletic, belonging in completely different genera or families. True Lepidotes were restricted down to just the original type speciesLepidotes gigas and a few of its closest relatives, all from the early Jurassic of Europe, while some other forms were moved into the newer genera Scheenstia and Callipurbeckia. Since then some other “Lepidotes” have also been reclassified, creating new names like Macrosemimimus, Occitanichthys, and Quasimodichthys.
There’s still work needing to be done on untangling all these Lepidotes-like fish – Scheenstia might actually now represent several different lineages, for example – but at least Lepidotes itself is now in a much better situation than it was just a couple of decades ago.
The ostrich-like “bird-mimic” dinosaur Ornithomimus was named in 1890, based on some hand and foot bones from Late Cretaceous-aged fossil beds in Colorado, USA.
The first ornithomimid known to science, it was initially thought to be a ornithopod, but then a few years later more fossil material revealed it was actually a theropod – and then it spent some time classified as a “megalosaur” before ornithomimids were finally recognized as being coelurosaurs in the early 20th century.
And for nearly a century after its discovery it was treated as a wastebasket taxon for any similar-looking fossil material from North America and Asia, with around 17 different species named within the genus. One of these was split off into Struthiomimus in 1917, but it wasn’t until much later that the rest began to get sorted out.
A review of known Ornithomimus fossils in the early 1970s renamed a couple more species into the new genera Archaeornithomimus and Dromiceiomimus, and dismissed most of the remaining species as dubious or invalid. Just two valid species now remained: the original Ornithomimus velox from Colorado, and Ornithomimus edmontonicus from Alberta, Canada.
Ornithomimus edmontonicus
Since then opinions have gone back and forth about some of the other Ornithomimus species. For a while Dromiceiomimus was merged back into Ornithomimus, but more recently it’s been found to have distinct limb proportions and was probably actually a separate genus after all. Another species that’s usually considered to be part of Struthiomimus is also sometimes instead classified as an Ornithomimus instead.
Really all of the North American ornithomimids are in need of a modern taxonomic revision – especially since Ornithomimus edmontonicus shows enough anatomical variation that it might actually represent a species complex of multiple very similar forms, which might get split apart in the future if anyone can figure out how to reliably distinguish them.
The rodent-like multituberculates were a major lineage of mammals that were only distantly related to modern marsupials and placentals. They originated around the time of the mid-Jurassic (~168 million years ago), survived through the end-Cretaceous mass extinction, and went on to become one of the most diverse and successful types of mammal in the Paleocene. After that point they began to decline, and after anw over-130-million-year-long run they went extinct* in the early Oligocene (~33 million years ago).
(* Except, possibly, in South America, where an enigmatic fossil known as Patagonia peregrina may represent a multi surviving as recently as about 18 million years ago in the early Miocene.)
First discovered in North America in the 1880s, Catopsalis foliatus was part of a group of multituberculates called taeniolabidoids. These multis got significantly larger than the rest of their kind – averaging beaver-sized but with some species getting up to at least capybara-sized – and were some of the first mammals to evolve into relatively big herbivores after the extinction of the non-avian dinosaurs.
Catopsalis foliatus
Catopsalis was named based on a partial jawbone and a few teeth, and over the next century or so various other similar-looking fossils from both North America and Asia were added into the genus as additional species. Eventually Catopsalis contained eight different species, ranging over about 10 million years from the late Cretaceous to the early Eocene – not especially big compared to some other wastebaskets we’ve looked at this month, but it was still a problem, muddying up attempts to understand the actual evolutionary relationships and biogeography of the taeniolabidoids.
Cladistic studies in the 1980s showed that Catopsalis was paraphyletic, made up of at least five separate lineages, and a few of them were subsequently renamed and reclassified. The Cretaceous Asian forms became Djadochtatherium and Catopsbaatar, and are now considered to be part of a different lineage of multis known as djadochtatherioids, while one of the remaining North American species then became Valenopsalis.
…But a couple of other new Catopsalis species have also been named in the meantime (one as recently as 2018), so there are still seven different species that need sorting out in this particular wastebasket.
Pseudosuchians, or “croc-line archosaurs”, are one of the two major lineages of archosaur reptiles, alongside the avemetatarsalians (pterosaurs and dinosaurs). Although today they’re represented only by crocodilians, they were especially successful and diverse back in the Triassic – and it was only after a mass extinction took out most of them that the dinosaurs were able to rise to prominence for the rest of the Mesozoic Era.
The first rauisuchians were discovered in the 1930s, represented only by fragmentary remains, and while they were initially recognized as being pseudosuchians their exact evolutionary relationships within that group were poorly understood for a long time. Over the next several decades they were classified with aetosaurs (early armored pseudosuchians), then ornithosuchids (even earlier pseudosuchians), and then erythrosuchids (not even pseudosuchians but an earlier type of archosauriform).
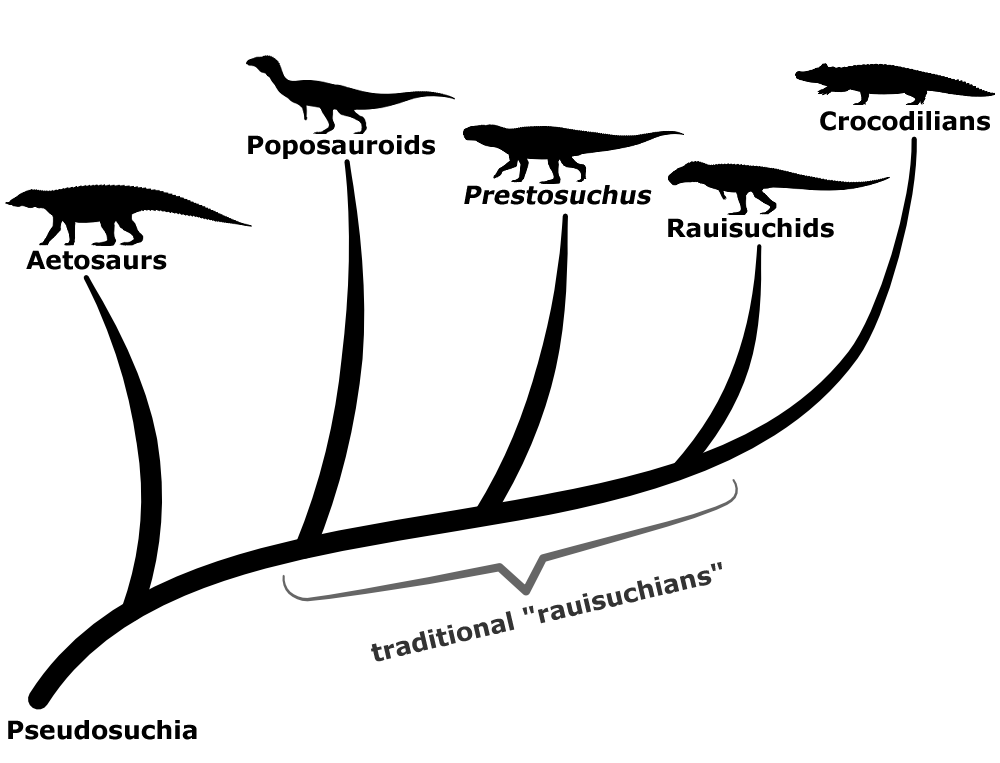
More complete fossil discoveries and better cladistic analysis methods in the 1980s led to them being classified as being very closely related to crocodylomorphs, with three main lineages recognized: the prestosuchids, the rauisuchids, and the poposauroids.
The “prestosuchid” Prestosuchus chiniquensis, the rauisuchid Postosuchus kirkpatricki, & the poposauroid Effigia okeeffeae (not to scale)
But even by the end of the 20th century “Rauisuchia” had never actually gotten a formal definition, and it had very much become a wastebasket taxon for a variety of paracrocodylomorph pseudosuchians that didn’t easily fit into any other major lineages.
In the 2000s renewed interest in rauisuchians’ anatomy and evolutionary relationships led to increasing recognition that they weren’t even a single defined group, with various species instead falling into different points along an “evolutionary grade“. The poposauroids and rauisuchids still seem to be distinct lineages, but the “prestosuchids” were found to be polyphyletic, with some forming a grade between the other two “rauisuchid” groups and others turning out to not even be paracrocodylomorphs.
And although the taxonomic concept of “Rauisuchia” as a distinct group has now been abandoned, the term “rauisuchians” does still remain in common use as an informal name for these animals – probably because it’s much more concise than saying “non-crocodylomorph paracrocodylomorphs”.
Protorthoptera was a group of fossil insects created in the early 20th century to categorize “primitive” neopterans – some of the earliest insects to have evolved the ability to fold their wings down over their backs. Known mostly from just fossilized forewings, they first appeared around 320 million years ago in the late Carboniferous, and after heavy losses during the Great Dying mass extinction they eventually disappeared in the mid-Triassic about 240 million years ago.
As early as the mid-20th century the protorthopterans were recognized as being a general taxonomic dumping ground, containing a mixture of early members of multiple different “orthopteroid” insect lineages. But invertebrate paleontologists at the time considered this collection of “primitive” insects to lack enough distinctive features to confidently separate them out from each other, and so the highly paraphyletic grouping continued to be used well into the 1990s.
Ctenoptilus elongatus
But in the early 2000s this situation finally changed. Proper cladistic analysis of protorthopteran fossils identified defining features of the wing vein patterns, and many species were reclassified into various lineages within the Archaeorthoptera – which includes modern grasshoppers, crickets, and locusts along with several closely related fossil groups like the titanopterans and caloneurodeans.
“Protorthoptera” is still sometimes used in a loose sense for fossil neopteran insects that still can’t be confidently classified anywhere else, so the wastebasket isn’t entirely cleared here.
And there are some alternate classification systems (mainly proposed by Russian paleontologists) that instead consider many protorthopterans to be notopterans closely related to modern ice-crawlers, and place others as part of other modern neopteran lineages such as webspinners and true bugs.
Hopefully better fossil discoveries and future studies will eventually help clear things up, and give us a better overall picture of the evolution of these insects.











{kind=link}