Mammalian tusks usually grow in symmetrical pairs with only minor developmental asymmetry, but a few species have evolved much more uneven arrangements.
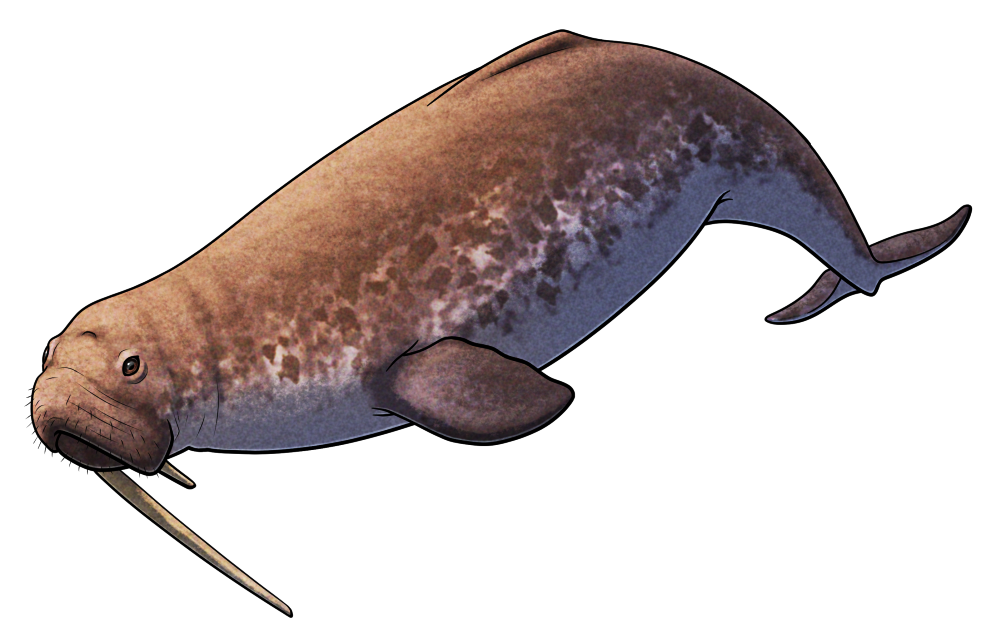
Odobenocetops peruvianus
Odobenocetops peruvianus was a small toothed whale that lived during the Miocene, about 7-3 million years ago, in shallow coastal waters around what is now Peru. Around 3m long (~10′), it was a highly unusual cetacean with binocular vision, a vestigial melon, muscular lips, and a pair of tusks – features convergent with walruses that suggest it had a similar lifestyle suction-feeding on seafloor molluscs and crustaceans.
In males the right tusk was much more elongated than the left, measuring around 50cm long (~1’8″) in this species and up to 1.35m (4’5″) in the closely related Odobenocetops leptodon. Since these teeth were quite fragile they probably weren’t used for any sort of combat, and they may have instead served more of a visual display function.
The woolly mammoth (Mammuthus primigenius) lived across Eurasia and North America during the last ice age, mostly from the Pleistocene about 400,000 years ago to the early Holocene about 10,000 years ago – altohugh a few relict populations survived until around 4,000 years ago in isolated areas of Alaska, Siberia, and eastern Russia.
Around 3m tall at the shoulder (~10ft), these hairy proboscideans had very long curving tusks that were used for digging out vegetation from under snow and ice, scraping bark from trees, and for fighting.
The tusks showed a lot of variation in their curvature, and were often rather asymmetrical, a condition also seen in the closely related Columbian mammoth. Like modern elephants mammoths may have also favored using one side over the other for certain tasks, which over their lifetimes could result in uneven wear exaggerating the natural asymmetry even more.
It was smaller than modern pronghorns, around 70cm tall at the shoulder (~2’4″), and males had long antler-like horns with three tines. Bizarrely, one of these horns was always at least twice the size of the other, with “left-horned” and “right-horned” individuals seeming to occur in equal numbers.
Meanwhile in China another Miocene ungulate known as Tsaidamotherium hedini also had strange headgear, with an enlarged right “horn” forming a helmet-like dome on top of its head. This species was featured here on the blog just year, so check out that post for more details about it.
Toothed whales – the branch of cetaceans that includes modern dolphins, porpoises, beaked whales, and sperm whales – have surprisingly asymmetrical skulls, with some of the bones skewed to one side and just the left nostril forming their blowhole.
Some of the most obvious external manifestation of this lopsidedness can be seen in sperm whales, which have their blowhole at the front left side of their head, and in male narwhals, which usually have a single left-side tusk.
This sort of asymmetry first appeared in the skulls of early toothed whales around 30 million years ago. And since the highest amounts of wonkiness have gone on to develop in lineages that hunt in dark, cluttered, or murky waters, this suggests that the trait is somehow linked to the evolution of complex echolocation.
Some ancient members of the river dolphin lineage also had some additional unusual asymmetry, sometimes having slightly sideways-bending snouts.
Ensidelphis riveroi was one of the weirdest of these, living around the coasts of what is now Peru during the Miocene, about 19 million years ago. Around 3m long (~10′), it had a very long narrow toothy snout that curved distinctly off to the right along its length.
Expectation vs reality
It’s not clear what the function of this bend was, or even if the only known skull actually represents the normal condition for this species. But Ensidelphis’ bendy snoot might have been used to probe around in muddy seafloor sediment or to extract prey from crevices, possibly like an underwater version of the modern wrybill.
Antaecetus aithai was an early whale that lived during the late Eocene (~40 million years ago) in what is now Morocco, at a time when northern Africa was covered by a warm shallow sea.
It was part of the “basilosaurids“, some of the first fully aquatic cetaceans – traditionally considered to be a single defined group, but more recently found to be more of an “evolutionary grade” of multiple early whale lineages – and much like Basilosaurus it had elongated back vertebrae that would have given it a very long slender body shape.
Antaecetus also had a proportionally smaller head and smaller teeth than other basilosaurids, along with much denser bones and a stiffer spine that would have made it a rather slow swimmer with reduced maneuverability. It was also fairly small overall compared to most of its relatives, probably around 6m long (~20′).
It was probably a slow-moving coastal water animal somewhat like modern sirenians – except unlike manatees and dugongs it was carnivorous. Its relatively delicate teeth suggest it was feeding on soft-bodied prey like cephalodpods, and with its lack of speed it must have been some sort of ambush predator, waiting around for potential prey to come within striking range.
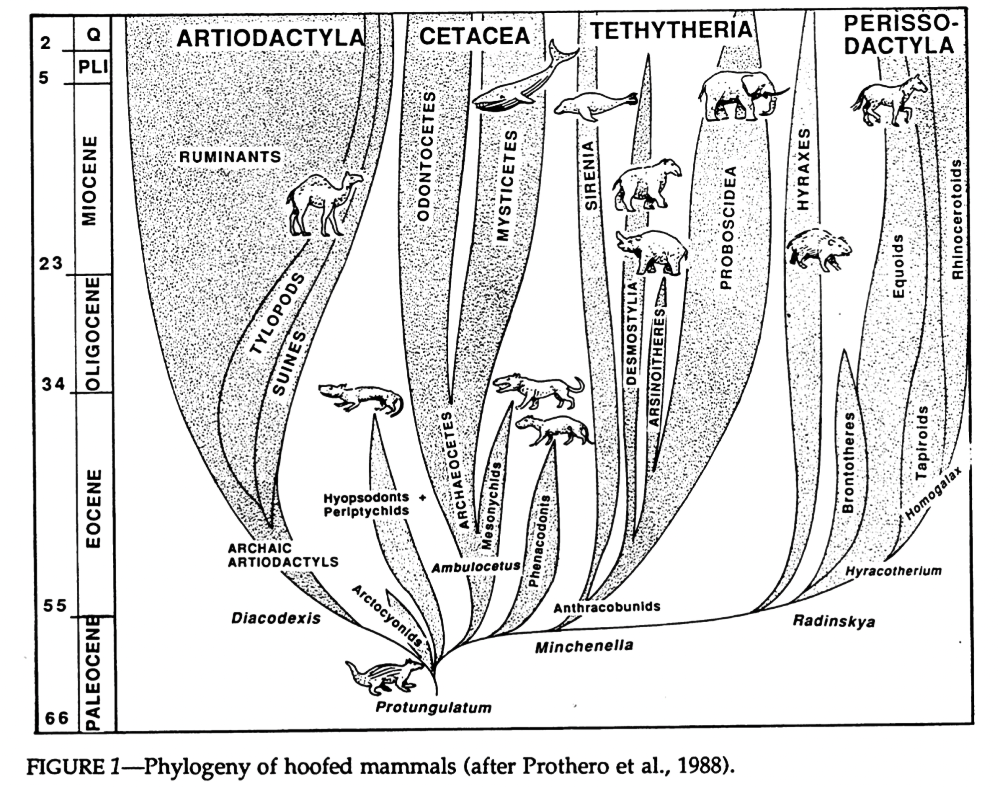
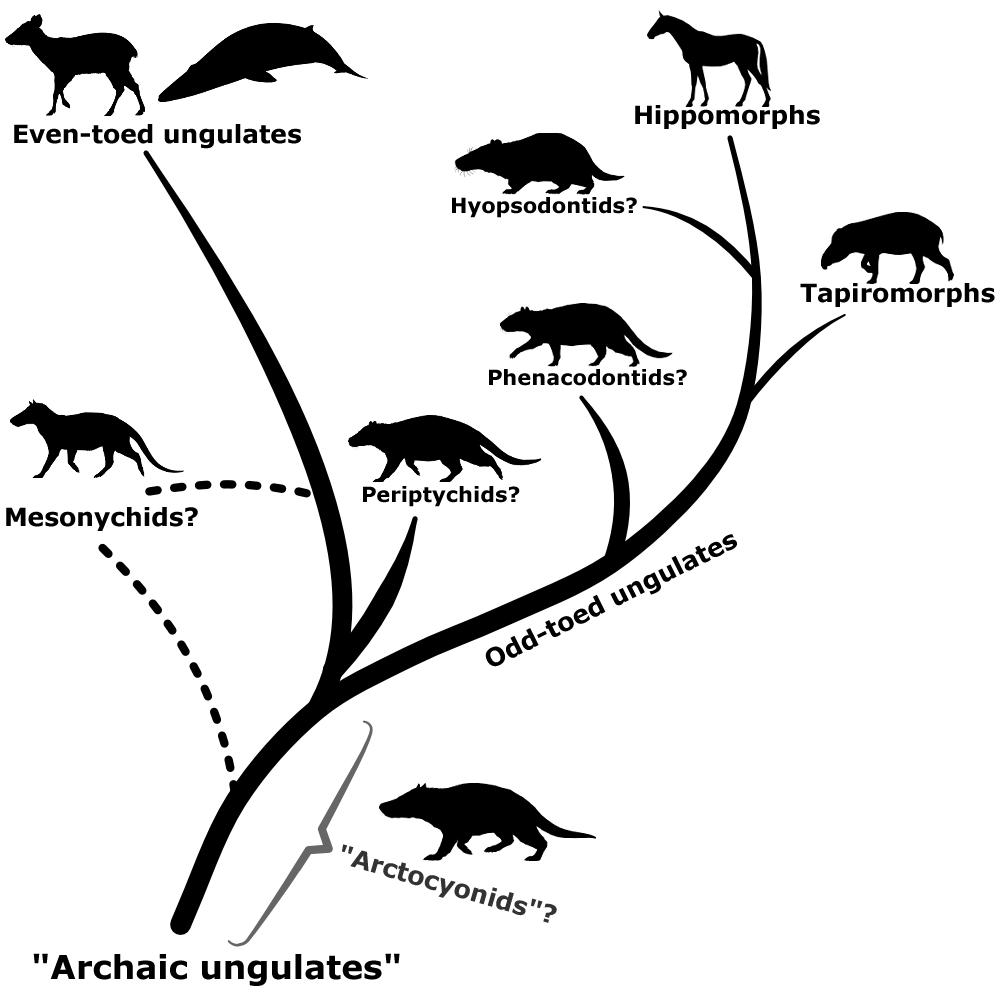
“Insectivora” was a wastebasket taxon so bad it had to be revised multiple times, but there’s another particularly infamous case in mammal taxonomy that’s still in the process of being resolved – the “condylarths“.
This group was first created in the early 1880s, during the Bone Wars, and initially was just a subgroup of odd-toed ungulates containing only the phenacodontids. But just a few years later Condylarthra was promoted up to its own order, and groups like the periptychids and hyopsodontids were added in too.
Then over the next few decades various groups were added and removed from the condylarths, most notably with the mesonychids and arctocyonids being brought in from their previous position with the creodonts.
By the mid-20th century the condylarths had become a big convenient dumping ground for any and all “primitive” ungulate-like mammals that didn’t easily fit into any modern groups, ranging in age from the early Paleocene through to the early Oligocene. But it soon became apparent that they had the same problem as the “insectivores” – there weren’t really any unique anatomical features that united all these animals together.
They generally had rounded-cusped molar teeth and hoof-like toes, but they also had rather generalized “primitive mammal” features and a diverse range of ecologies. Some were small herbivores, but others were coati-like or dog-like omnivores, and some were even bear-sized carnivores.
From left to right, top row: Hyopsodus lepidus (hyopsodontid), Meniscotherium chamense (phenacodontid), Arctocyon primaevus (“arctocyonid”). Bottom row: Ectoconus ditrigonus (periptychid), Mesonyx obtusidens (mesonychid)
It wasn’t even clear how the various different condylarth groups were actually related to each other. The best guess was that arctocyonids had arisen from within the “insectivores”, with a Protungulatum-like form as the common ancestor of all the other condylarths. Where exactly modern ungulates had then evolved from within the condylarths was also still uncertain.
Cladistic analysis in the 1980s began to tackle the confusing pile of assorted condylarths, and showed that they weren’t the single ancestral source of all modern ungulates, but instead a loose collection of several unrelated groups from all over the ungulate evolutionary tree. Arctocyonids, periptychids, and hyopsodontids were placed as early “primitive” lineages, phenacodontids were loosely linked with the ancestors of odd-toed ungulates once again, and mesonychids were considered to be the ancestors of whales.
And, once again paralleling the mess of the “insectivores”, it wasn’t until genetic methods became available in the late 1990s that larger-scale ungulate relationships began to be properly resolved. The paenungulates (elephants, hyraxes, and sirenians), which had been traditionally considered to be a major branch of ungulates, were removed entirely and reclassified as afrotheres. And, along with some new fossil discoveries, whales were recognized as having actually evolved from within the even-toed ungulates instead of from mesonychids.
This shake-up threw the still-problematic “condylarth” classifications back into question – with some “condylarths” turning out toalso be afrotheres instead of true ungulates.
Today the actual relationships of the main “condylarth” ungulate families are still in the process of being figured out, and there’s a lot of remaining uncertainty and disagreement about them.
Phenacodontids seem to have mostly maintained their traditional position as early odd-toed ungulates, and hyopsodontids may potentially be part of this group too – possibly as members of the hippomorph lineage, closely related to horses and brontotheres. Arctocyonids might be a wastebasket themselves, with some studies finding them to be a mix of several different archaic ungulate lineages. Periptychids may have links to the even-toed ungulates. The mesonychids, meanwhile, are now generally considered to be a separate order from the traditional “condylarths”, and may be either an early branch of the even-toed ungulates or much more basal ungulates closely related to the “arctocyonids”.
Since the term “condylarth” no longer has any real taxonomic meaning some paleontologists have proposed replacing it with “archaic ungulate” to distance from the historical messiness of the old name. But this hasn’t really caught on, and many papers still use “condylarth” in a very loose sense to refer to an “evolutionary grade” of early ungulates of unclear evolutionary affinities.
And while that’s the last main entry for this month, we’re not quite done yet. There’s still one weekday left in October, and after digging through so many taxonomic garbage cans there’s only one place we can go now.
Cetotheres were a group of small baleen whales, one of three major lineages of these cetaceans alongside the rorquals and the right whales. They first appeared in the fossil record in the mid-Miocene, about 14 million years ago (but are estimated to have actually originated 10-15 million years earlier), and disappeared during the Pleistocene about 2 million years ago.
First recognized in the mid-19th century, for a long time the cetotheres were used as a wastebasket for all fossil baleen whales that didn’t clearly fit into any modern whale families. By the start of the 21st century nearly 30 different genera representing numerous different species were all lumped into the group – and the genus Cetotherium was another wastebasket in itself with at least 12 assigned species, many of which were based on fragmentary or dubious remains.
Ciuciulea davidi
This was finally cleaned up in the 2000s, when a revision of the cetotheres cut the group down to just 6 genera. Since then a handful of additional new genera and species have been named, and while a few polyphyleticCetotherium species may still need tidying up the cetotheres have overall gone from being a total taxonomic mess to actually being one of the best studied groups of fossil baleen whales.
Their exact evolutionary relationships with each other are still in flux, but the most surprising discovery from the improved understanding of these ancient whales is that they might not be extinct after all.
At this point it seems fairly likely that the pygmy right whale really is either the last surviving representative of the cetothere lineage, or at least is a very close evolutionary “cousin” (a “cetotherioid”) closer related to them than to any other modern baleen whales.
Tsaidamotherium hedini was a ruminant ungulate living around 11 million years ago during the late Miocene, in the northeastern part of the Tibetan Plateau in what is now Northwestern China. Although it’s known only from partial skull remains it was probably similar in body size to a large sheep, about 80cm tall at the shoulder (2’7″).
Since its discovery in the 1930s it’s traditionally been classified as part of the muskox lineage, but in 2022 it was proposed to actually be a giraffoid very closely related to the newly-discovered Discokeryx.
Tsaidamotherium had some extremely unusual headgear, with highly asymmetrical “horns” (actually ossicones if was a giraffoid). The left one was small and positioned above the eye, while the right one was shifted back and towards the middle of the forehead, and was expanded out into a wide bony disk that would have supported a large helmet-like domed keratin covering.
Its skull also had a very large nasal cavity resembling that of the modern saiga antelope, suggesting it may have convergently evolved a similar sort of complex air-filtering snout to deal with dry cold air in its mountainous habitat.
Modern beluga whales and narwhals are the only living representatives of the monodontid lineage, found only in cold Arctic and sub-Arctic waters. But this whale family actually first evolved in much warmer climates – and some of them were downright tropical.
It seems to have had a larger number of functional teeth than modern monodontids, and probably didn’t suction feed like its modern close relatives. Instead it may have fed more like most porpoises and dolphins, relying more on speed and snapping jaws to capture prey.
It inhabited the Mediterranean at a time not long after the sea there had mostly dried up and then been rapidly refilled. The presence of warm-water marine species such as bull sharks, tiger sharks, and dugongs in the same fossil beds as Casatia indicates the local climate at the time was hotter than it is today, with tropical temperatures – and suggests that this whale’s ancestors must have originally moved into the replenishing Mediterranean from lower latitudes alongside these other warmth-adapted animals.
This tropical monodontid was also much closer related to modern belugas than modern narwhals are, which raises the possibility that the two living monodontid species actually specialized for colder conditions completely independently of each other rather than descending from a cold-adapted common ancestor. Instead modern belugas and narwhals may have originated from separate warm-water monodontid ancestors who evolved similar cold-tolerant adaptations in parallel as the climate cooled during the onset of the Quaternary ice age, while the rest of their relatives all went extinct.
Known just from fossilized lower jaws and teeth, with some teeth up to nearly 13cm long (~5″), its full life appearance and size are uncertain – but it may have been slightly larger than a modern bottlenose dolphin at around 4.5m long (~14’9″). It’s traditionally been considered to be part of the kogiid family, closely related to modern pygmy and dwarf sperm whales, but some studies disagree with that classification and instead place it in the true sperm whale lineage.
It was probably a predator in a similar ecological role to modern orcas, adapted for hunting prey like squid, fish, and smaller marine mammals. But unlike orcas it wouldn’t have been the apex predator of its ecosystem, subject to predation pressure by even larger carnivores like macroraptorial sperm whales and everyone’s favorite ridiculously huge shark – and as a result it probably had a “live fast and die young” lifestyle similar to modern kogiids and other small-to-medium-sized Miocene physeteroids, rapidly maturing and only living to around 20 years old.
I’ve reconstructed Kogiopsis here as a kogiid-like animal, with a similar sort of shark-like head shape and “false gill” markings. In the background a second individual is depicted displaying “inking” behavior, releasing a defensive cloud of reddish-brown fluid from a specialized sac in its colon.
Living during the mid-Eocene, about 43 million years ago, in a shallow sea-covered region that is now part of Egypt‘s Western Desert, Phiomicetus was an early protocetid – an amphibious foot-powered swimmer, at a transitional point in the evolution of whales from deer-like terrestrial animals to fully aquatic screaming torpedoes.
About 3m long (~10′), it had large jaw muscles and sharp teeth with wear patterns that suggest it was a raptorial hunter grabbing and snapping at prey with powerful bites. It would have probably tackled fairly big prey compared to other protocetids, hunting things like large fish, turtles, and even smaller whales in an ecological role similar to that of modern orcas.
Along with the distantly-related long-snouted Rayanistes it’s one of the earliest known whales from Africa, giving us further glimpses at a time period when early cetaceans were first dispersing out from the South Asian subcontinent via the ancient Tethys Sea.








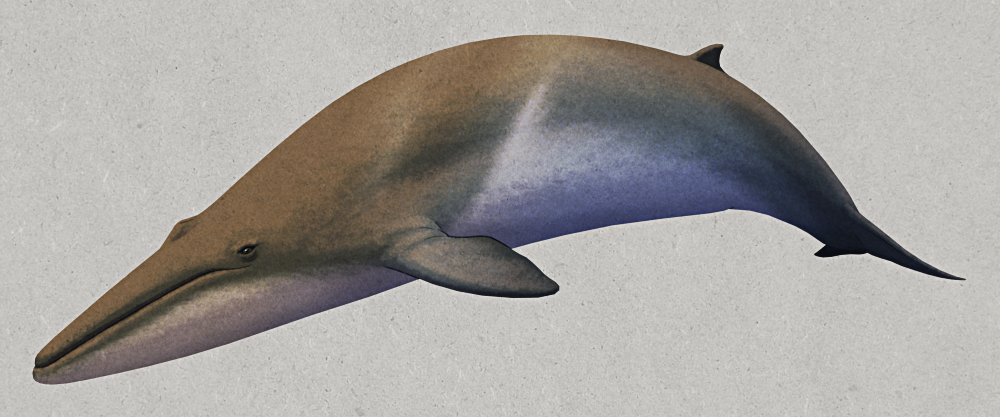





{kind=link}
{kind=link}
{kind=link}