

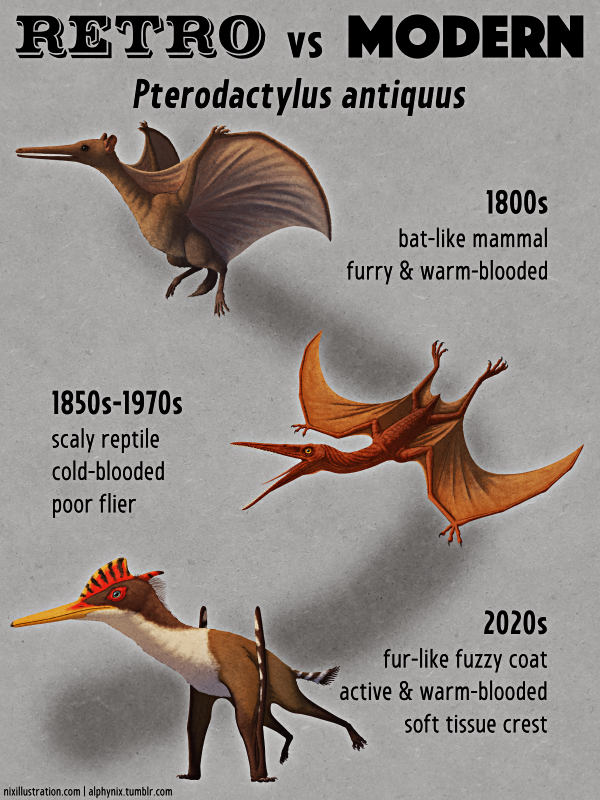
Deinonychus antirrhopus was one of the most significant dinosaur discoveries of the 20th century, kicking off the Dinosaur Renaissance and the recognition of the evolutionary link between maniraptoran theropods and modern birds.
1960s
The first remains of this species were discovered in North America in the 1930s, but at the time the fossils weren’t officially described or named. It wasn’t until the 1960s that more specimens were found in Montana, representing at least three preserved individuals, and paleontologist John Ostrom recognized that there was something very special about this dinosaur.
In contrast to the prevailing view at the time that theropods were all upright tail-dragging “sluggish lizards” this was clearly a highly specialized and active predator, with a huge sickle-shaped claw on each foot and a long stiff tail for balance – inspiring its scientific name’s meaning of “counterbalancing terrible claw”.
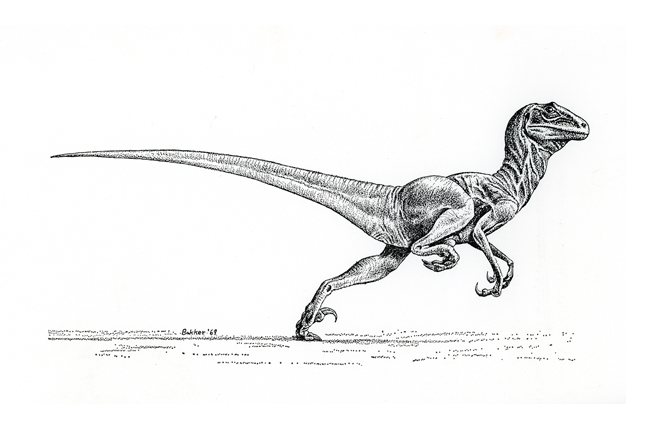
And while the very first reconstruction of Deinonychus might seem retro to modern eyes, at the time it was revolutionary and it went on to become an iconic representation of the species for the next couple of decades. Drawn by Robert Bakker, who was Ostrom’s student at the time, it depicted a lizard-like creature with its body held in a horizontal pose and its tail held out straight behind it. Its head was portrayed as more domed than we now know Deinonychus’ skull to have been, and its neck was up in an alert posture while the animal ran at full sprint, with its sickle-claws held up away from the ground to keep them sharp.
A few years later further discoveries showed a highly bird-like pelvis and hands very similar to those of Archaeopteryx, triggering the Dinosaur Renaissance reinterpretation of dinosaurs as active warm-blooded animals, and the revival of the 19th century idea that they were the ancestors of birds.
1980s-1990s
As the “birds are dinosaurs” idea began to gain acceptance with increasing amounts of anatomical evidence, some paleontologists in the 1980s began to also suspect that highly bird-like dromaeosaurids like Deinonychus might have also been feathered. Some reconstructions during this time showed this to varying degrees, particularly those drawn by Bakker and by Gregory Paul – but it didn’t really catch on more widely at first, for one very big reason:
Jurassic Park happened.
Dromaeosaurs hadn’t been well-known dinosaurs to the general public before that point, but the 1993 JP “raptors” were an instant hit in pop culture. Physically based much more on Deinonychus than on Velociraptor, and exemplifying the renaissance view of dinosaurs in major media for the first time, the movie’s fully scaled and oversized version of these animals dominated popular depictions for years afterwards. Even the most rigorous and anatomically accurate artwork showcasing their bird-like features still usually kept them completely naked to retain that familiar reptilian appearance.
Most 1990s attempts at any feathering tended towards being as sparse as possible – often along with the shrinkwrapping typical for the era – at best being decent for the time but what we’d now deride as “half-assed”, and at worst being “a few token quills on the back of the head”.
Deinonychus fossils found in association with Tenontosaurus were also interpreted as being evidence of cooperative pack hunting behavior during this time, and it became a common paleoart meme to depict the large herbivore being constantly swarmed by ravenous raptors.
2020s
The mid-1990s discovery of fully-feathered dinosaurs like Sinosauropteryx in China, followed a few years later by raptors with wing-feathered arms like Sinornithosaurus, gradually began to put the fluff back onto dinosaurs like Deinonychus.
(…At least in reasonably scientific paleoart. The much much stronger and ongoing resistance from popular culture is far too big a subject to get into here. But maybe, just maybe, we’re finally hitting a turning point there?)
Early attempts at properly feathering dinosaurs were a bit awkward, usually looking rather like a bunch of scruffy greasy hair glued onto a scaly raptor, a dinosaur wearing fuzzy pajamas, or like the old “bird-lizard” depictions of Archaeopteryx. Even into the early 2010s some paleoart memes were still common in depictions of dromaeosaurs, but increasingly better understanding of their anatomy and plumage arrangements over the last decade or so has brought us to a much more birdlike interpretation of these animals – with paleoartists like Emily Willoughby being especially influential in popularizing the modern view of dinosaurs like Deinonychus.
We now know Deinonychus lived during the Early Cretaceous, about 115-108 million years ago, in what is now the Mountain West and South Central United States. Up to around 3.4m long (11′), it stood about 1m tall (3’3″), similar in size to a large dog.
It had blade-like teeth in its jaws, and forward-facing eyes with stereoscopic vision. Its three-fingered arms would have been covered by wing-like feathers, and its tail probably had feathers all the way along its length and was stiffened but not totally inflexible.
It may have used the sickle-claws on its feet to pin down struggling prey, eating it alive while flapping its wings and waving its tail for balance. And while often depicted as an extremely fast-runner, its leg proportions and foot anatomy suggest it was actually built more for walking and had an especially strong grip strength in its feet, trading speed for power and probably being more of an ambush predator – often being compared to a “giant ground-hawk”.
Pack hunting has been called into question recently, too, arguing that the Tenontosaurus sites may actually represent crocodile-like or Komodo dragon-like behavior with mobs of scavenging individuals congregating at a carcass. But other evidence from trackways and Utahraptor does offer potential support for pack behavior in raptors, so it’s still open to interpretation.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}