I’m still trying to work through that big pile of speculative evolution concepts from a few years ago, so I’m hoping to make this month sort of a “lightning round” to finally clear out the backlog.
(I’m not going to set a definite posting schedule this year because things are pretty chaotic right now. But I’ll try to fit in as many as I can!)
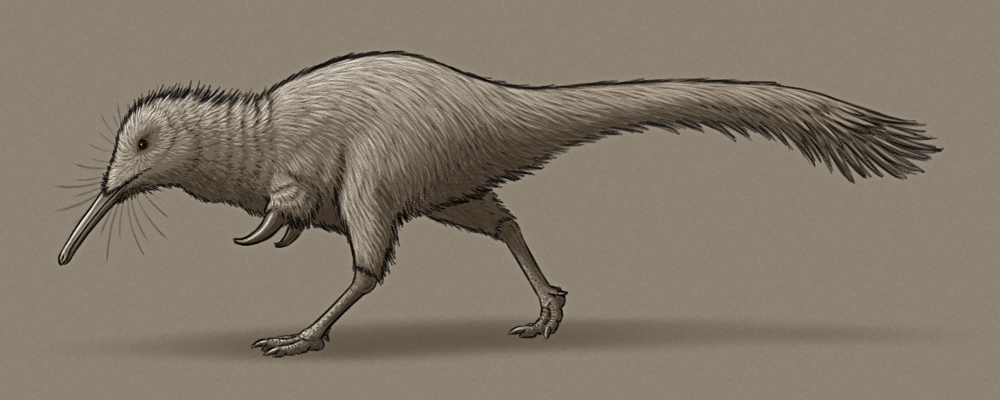
So let’s start off with a concept from an anonymous submitter, who requested a “kiwi/sengi niche alverezsaur”:
Khamartaia dolabella is similar in size and build to Shuvuuia, about 1m in length (3’3″), with slender legs and stumpy arms with massive thumb claws. Unlike its close relatives, however, it has small eyes and fairly poor vision, relying more on its other senses to forage around during the darkness of night.
It has an acute sense of smell, and its long narrow snout is full of highly touch-sensitive nerves, allowing it to probe around for invertebrate prey in soil, undergrowth, and cracks and crevices. Its chunky thumb claws are used to dig up burrows and to tear through bark to access deeper insect nests.
It mainly relies on its long legs to sprint away from threats, although with its poor eyesight these escapes are often rather ungainly.
Although it’s not visible externally, owls have one of the most striking modern examples of asymmetry. The ears of many species are uneven, with the right ear opening positioned higher up than the left, giving them the ability to pinpoint the sounds of their prey much more accurately.
But surprisingly this isn’t a unique anatomical trait that only ever evolved once in their common ancestor.
Instead, multiple different lineages of owls have actually convergently evolved wonky ears somewhere between four and seven separate times.
The boreal owl (Aegolius funereus), also known as Tengmalm’s owl, is a small 25cm long (~10″) true owl found across much of the northern parts of both Eurasia and North America. While most other owls’ asymmetrical ear openings are formed just by soft tissue, the boreal owl’s lopsided ears are actually visible in the bones of its skull.
But despite how many times owls have convergently evolved asymmetrical ears, and how successful this adaptation has been for them, for a long time it seemed to be something that no other animals have ever mimicked.
In the early 2000s asymmetric ears were reported in the skulls of some troodontid dinosaurs, which seem to have been nocturnal hearing-based hunters similar to owls, but proper details on this feature still haven’t been formally published.
The night parrot (Pezoporus occidentalis) is a small ground-dwelling parrot found in Australia, close to the same size as the boreal owl at around 22cm long (~9″). Critically endangered and very elusive, it’s rarely seen and little is known about it – and it was presumed extinct for much of the 20th century, until more recent sightings of living individuals confirmed that the species is still hanging on.
Recent studies of preserved museum specimens have revealed that it seems to have poor night vision but excellent hearing, and that its right ear opening is noticeably asymmetrical, bulging out sideways from its skull. Much like owls the night parrot relies on acute directional hearing to navigate in darkness, but since its diet consists mainly of seeds it’s probably not using this ability to locate food sources. Instead it may be listening out to keep track of the precise locations of other parrots, and for the approach of predators – so its sharp sense of hearing may be the reason this unique bird has so far just barely managed to survive the presence of invasive cats and foxes.
Many decapod crustaceans have slightly asymmetrical pincers, often with one claw being chunkier and specialized for “crushing” while the other is more slender and used for “cutting”.
But fiddler crabs take this sort of asymmetry to the extreme as part of their sexual dimorphism – males have one massively oversized claw, which is used for both visual display to potential mates and for physical fights against rivals.
Some of the earliest fiddler crabs are known from the Miocene of what is now northern Brazil. Although the fossils have been given several different taxonomic names since their discovery in the 1970s (including Uca maracoani antiqua, Uca antiqua, and Uca inaciobritoi) they’re currently considered to be indistinguishable from the modern Brazilian fiddler crab, Uca maracoani, meaning that these crabs have remained externally unchanged for the last 16 million years.
Up to about 4cm in carapace width (~1.6″), modern Uca maracoani are found in coastal mangrove swamps and tidal mudflats around the northern and eastern coasts of South America – and some of these environments have also undergone little change since the Miocene. Males of the species can develop their enlarged pincer on either side of their bodies, with lefties and righties seeming to occur in equal numbers.
Most of the wastebasket taxa featured this month are completely extinct and known only from fossils, but to start things off let’s take a look at a major example of how even groups with living members could have their classification muddled up for centuries.
The name Insectivora first came into use in the early 1820s, and was used to refer to various “primitive-looking” small insect-eating mammals, with modern shrews, moles, hedgehogs, tenrecs, and golden moles as the original core members.
By the early 20th century insectivorans were considered to represent the “primitive” ancestral stock that all other placental mammals had ultimately descended from, and any vaguely similar fossil species also got dumped under the label. Extinct groups like leptictids, cimolestans, adapisoriculids, and apatemyids all went into the increasingly bloated Insectivora, too, making the situation even more of a wastebasket as time went on.
…Insectivora?
The problem was that the only characteristics that really united these various animals were very generic “early placental mammal” traits – small body size, five clawed digits on the hands and feet, relatively unspecialized teeth, and mostly-insectivorous diets – and attempts at making sense of their evolutionary relationships were increasingly convoluted.
The rise of cladistic methods from the 1970s onwards resulted in a lot of “insectivores” finally being recognized as unrelated to each other, removing them from the group and paring things back down closer to the name’s original definition. The idea that insectivorans were ancestral to all other placentals was abandoned, instead reclassifying them as being related to carnivorans, and the remaining members were recognized as just retaining a superficially “primitive” mammalian body plan.
Just shrews, moles, hedgehogs, solenodons, tenrecs, and golden moles were left, and to disassociate from the massive mess that had been Insectivora this version of the group was instead now called Lipotyphla.
Lipotyphla
But there were still no unique anatomical links between the remaining lipotyphlans. And then once genetic methods became available in the late 1990s, something unexpected happened.
Lipotyphla was suddenly split in half. For a while it was unclear if even the remaining shrew-mole-hedgehog-solenodon group was still valid – hedgehogs’ relationships were especially unstable in some studies – but by the mid-2000s things began to settle down into their current state.
Finally, after almost 200 years of confusion, the insectivore wastebasket has (hopefully) now been cleaned up. The remaining “true lipotyphlans” do seem to all be part of a single lineage, united by their genetics rather than by anatomical features, and are now known as Eulipotyphla.
A few fossil groups like nyctitheriids and amphilemurids are generally also still included, but since this classification is based just on their anatomy it isn’t entirely certain. The only exception to this are the nesophontids, which went extinct recently enough that we’ve actually recovered ancient DNA from them and confirmed they were eulipotyphlans closely related to solenodons.
Squeezing in one last bonus #Spectember post this year!
This one isn’t based on a specific prompt, but instead is a companion piece to a previous one.
While North American brontotheres were adapting to the spread of grasslands, some of their Asian cousins took a very different evolutionary path through the rest of the Cenozoic.
Today’s #Spectember concepts come from three submitters: anonymous, Jonas Werpachowski, and Novaraptoria.
Humdertaker (Suchomergus pollinctor)
Despite having a convergent resemblance to penguins or gannetwhales, the humdertaker (Suchomergus pollinctor) is actually a distant descendant of modern hummingbirds.
Today’s #Spectember concept is a combination of a couple of anonymous submissions:
Crowned brontothere (left) and woolly paracerathere (right)
These two animals are the descendants of brontotheres and paraceratheres, almost the last living representatives of their kinds, hanging on in the equivalent of modern-day times in a world similar to our own.
Despite some minor delays, it’s time once again for #Spectember – when I dive back into the big pile of speculative evolution concepts that you all submitted to me in 2020, and try to get through a few more of the backlog.
(…There’s still over 50 of them left. This is going to take a while.)
So today’s concept comes from an anonymous submitter, who requested an arboreal ornithopod dinosaur:
Most trilobites were able to roll themselves up into a protective ball – a behavior known as enrollment or volvation – exposing just their heavily armored backs to attackers. They’re often found fossilized curled up like this, and rare preservation of soft tissues shows that they had a complex system of muscles to help them quickly achieve this pose while simultaneously tucking their antennae and all their limbs safely inside their enrolled shells.
Some species also developed sharp defensive spines and spikes that jutted out when they enrolled, making themselves even more daunting to potential predators in one of the earliest known examples of an evolutionary “arms race”.
The major groups of the euarthropods are the chelicerates, mandibulates, and the extinct artiopodans, but there were some Cambrian species that still can’t be easily fitted in to any of those lineages.









{kind=link}