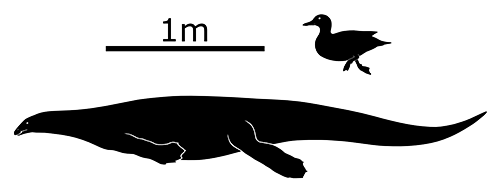
Atopodentatus unicus lived in Southwest China during the early-to-mid Triassic, around 247-240 million years ago. About 3m long (9’10”), it was a marine reptile – probably part of an early branch of the sauropterygians – with an elongated streamlined body and paddle-like limbs.
When it was originally described in 2014 it seemed to have a head unlike anything seen before. The skull of the only known fossil specimen was incomplete and badly crushed, but it was reconstructed as having a downward-hooking upper jaw with a vertical split in the middle forming a zipper-like row of teeth.
The original version of Atopodentatus
But then just two years later some more complete skulls were discovered and revealed something completely different: the projections on Atopodentatus‘ snout actually stuck out to each side in a wide flat “hammerhead” shape on both its upper and lower jaws.
Not quite as bizarre as before, but still a Triassic weirdo!
It also seems to have been a rare example of a herbivorousMesozoic marine reptile, probably rooting around on the seafloor with its shovel-like mouth, using its chisel-shaped front teeth to scoop up mouthfuls of algae and other marine plants and then straining out the water through its closely spaced needle-like back teeth.
We can’t go through this month without having an appearance from the most famous group of weird-headed dinosaurs: the ceratopsids!
Their distinctive-looking skulls were highly modified from those of their ancestors, with large bony frills extending from the back of their heads, various elaborate horns and spikes, enormous nasal cavities, large hooked beaks at the front of their snouts, and rows of slicing teeth further back.
And while typically depicted as purely herbivorous, ceratopsids’ powerful parrot-like beaks and lack of grinding teeth suggest they may actually have been somewhat more omnivorous – the Cretaceous equivalent of pigs – still feeding mainly on plant matter but also munching on carrion and opportunistically eating smaller animals when they got the chance.
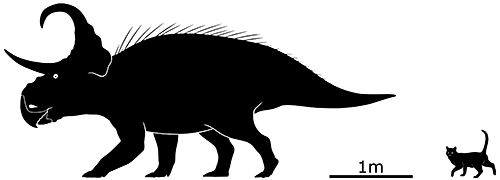
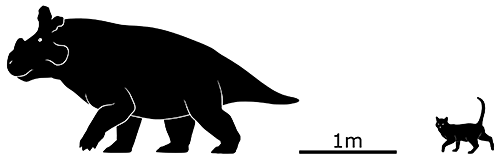
Machairoceratops cronusi here lived during the late Cretaceous of Utah, USA, about 77 million years ago. Only one partial skull has ever been found belonging to an individual about 4.5m long (14’9″), but it wasn’t fully grown and so probably reached slightly larger sizes.
It had two long spikes at the top of its frill, similar to its close relative Diabloceratops but curving dramatically forward and downwards above its face. Whether they were purely for display or used in horn-locking shoving matches is unknown, but either way it was a unique arrangement compared to all other known ceratopsids.
The dinoceratans were a lineage of hoofed herbivorous mammals whose evolutionary affinities are a little uncertain, but may have been related to the South American meridiungulates. Found in Asia and North America from the late Paleocene to the late Eocene, they had bulky rhino-like bodies and were some of the largest terrestrial animals of their time.
Eobasileus cornutus was one of the biggest of them all, measuring around 2.1m tall at the shoulder (~7′) and living in the Western United States during the early Eocene, about 46-40 million years ago.
And it had a very odd-looking head, with six blunt ossicone-like horns, large sabre-like fangs, bony flanges on its lower jaw, a concave forehead, and a proportionally tiny brain for its body size. The horns and fangs were sexually dimorphic, much smaller in females, suggesting they were mainly used for display or combat between males.
Dilophosaurus wetherilli is a fairly recognizable dinosaur thanks to its memorable appearance in the Jurassic Park franchise – but unfortunately that also means the popular image of it is completely wrong.
Rather than a small frill-necked venom-spitting creature, this early theropod was actually rather large, reaching around 7m long (~23′), and along with its distinctive double crests it also had a narrow snout with large teeth and a distinctive notch at the front of its lower jaw.
It lived in North America during the early Jurassic, about 196-183 million years ago, and while it wasn’t venomous its notched jaws were probably capable of delivering powerful bites to small struggling prey, much like the similar-looking ornithosuchids in the Triassic. Some structural similarities to the skulls of spinosaurids suggest it may have primarily eaten fish.
Its two bony crests were probably used for visual display, with juveniles only having small crests that fully developed as they matured. They also may have had a more extensive keratinous covering, so it’s not clear what their actual shape and full extent was in life.
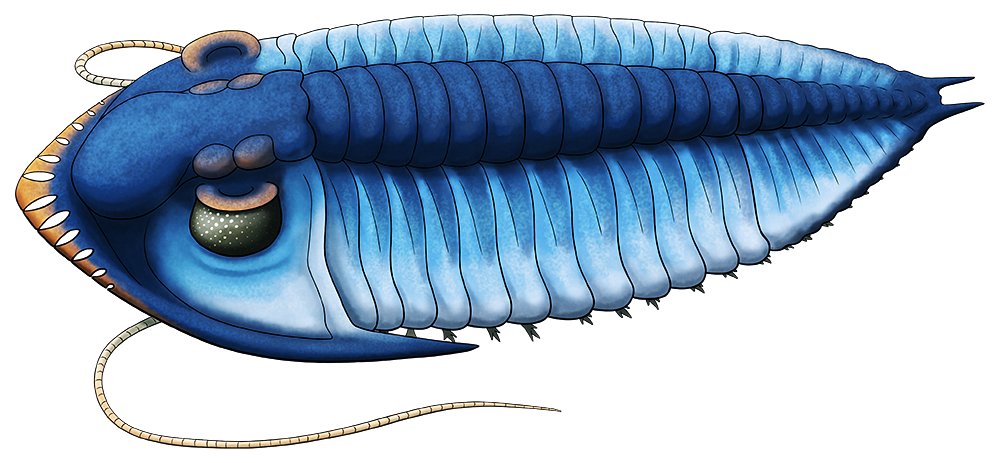
The eugeneodontidans were a group of cartilaginous fish which convergently evolved to resemble sharks but were much closer related to modern chimaeras. Due to their cartilage skeletons usually little more than their teeth are found as fossils, and for a long time their ecology and life appearance has been poorly understood because of just how weird those teeth were.
These fish had unique “tooth whorls” in their lower jaws, and the most famous member of the group is probably Helicoprion, with the exact anatomical placement of its buzzsaw-whorl only being properly figured out in 2013.
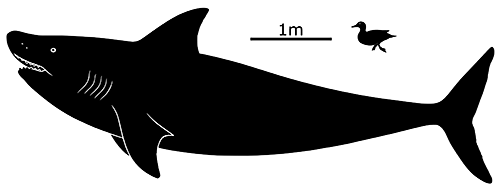
But another eugeneodontidan named Edestus was equally strange.
Living during the late Carboniferous, about 306-299 million years ago, Edestus giganteus was the largest species in the genus, reaching estimated lengths of up to 6m (19’8″), similar in size to a modern orca or a particularly large white shark.
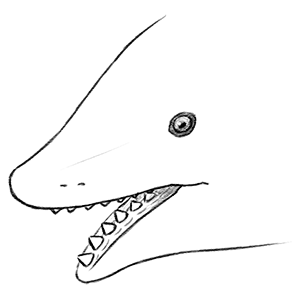
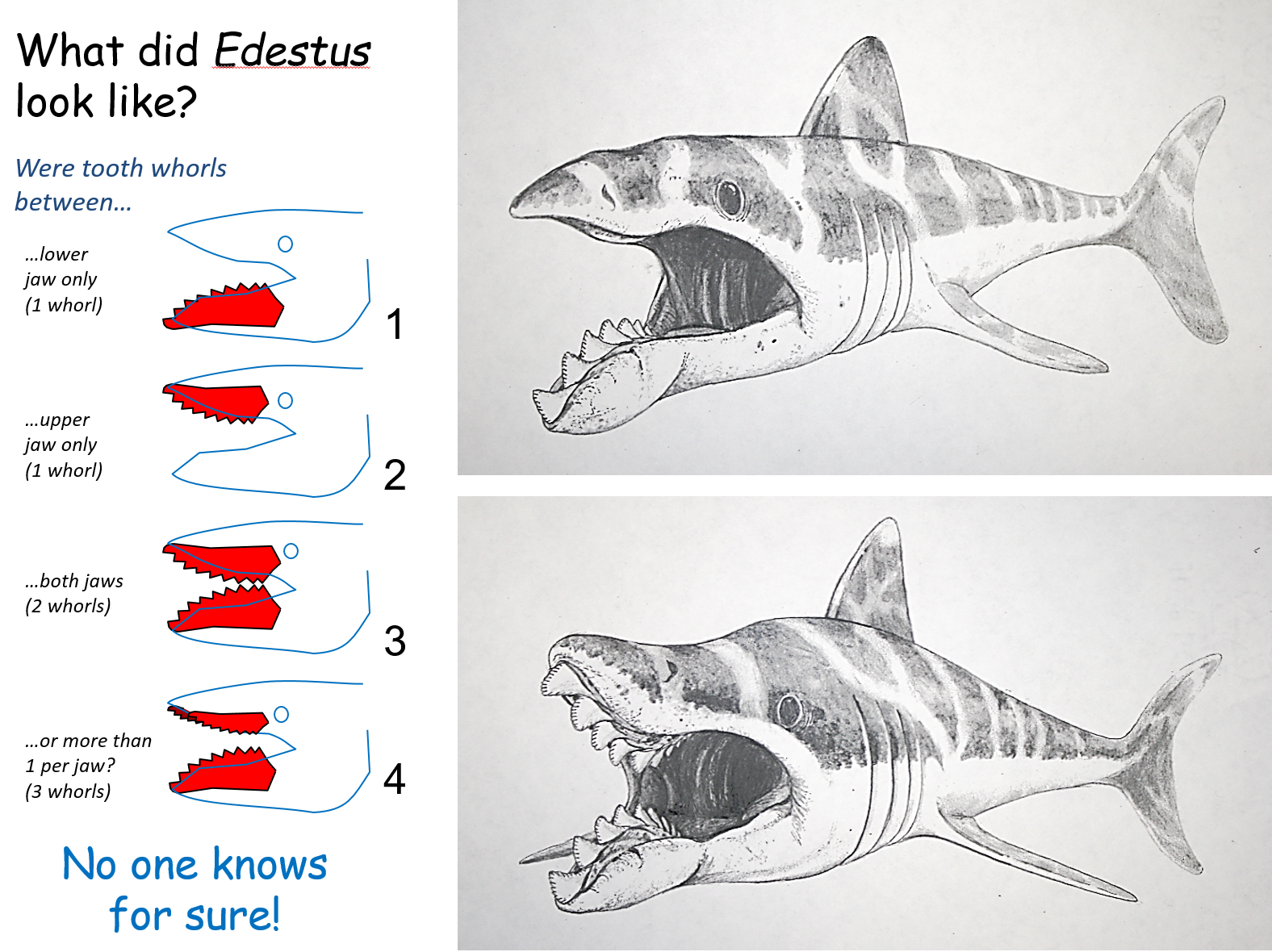
Let’s take a closer peek at that mouth.
Yes, that’s a single central row of teeth in both its upper and lower jaws.
Edestus‘ whorls grew in curving “banana-shaped” brackets that resembled an enormous pair of pinking shears, with new teeth being added on at the back and the oldest teeth occasionally being ejected off from the front. How this jaw arrangement worked was a longstanding paleontological mystery, with variousbizarreideas being proposed over the years – until a particularly well-preserved skull was analyzed in early 2019, revealing a two-jointed system in its lower jaw that allowed it to move its tooth brackets quickly back and forth, using a “snap-and-slice” motion to grab hold of prey like fish and soft-bodied cephalopods and cut them in half.
Along with body impressions from other related eugeneodontidans like Fadenia, showing a shark-like tail and a complete lack of rear fins, we now have a much better picture of what this bizarre fish probably looked like.
A little more closely related to modern mammals, the dinocephalians were a a diverse group that were found across Pangaea during the middle of the Permian period. Many of them had thickened skulls that may have been used for headbutting each other, and some also developed bony horn-like projections around their faces.
And Estemmenosuchus mirabilis here was particularly elaborately ornamented, earning it a name meaning “wondrous crowned crocodile”. It lived in the Perm region of Russia during the mid Permian, about 268-265 million years ago, and was one of the largest dinocephalians, reaching at least 3m long (9’10”).
It had two big antler-like structures on its head, two wide cheek flanges, and a small nose horn, almost looking like the synapsid version of a ceratopsid dinosaur – and with its big bulky body, fairly erect-legged posture, and herbivorous-or-omnivorous diet it may have been a fairly close ecological equivalent to them, too.
But it’s also possible it was semi-aquatic, and it certainly does have a very hippo-like appearance when reconstructed with a decent amount of soft tissue.
One specimen of Estemmenosuchus even preserved skin impressions around its face, which were described in Russian in the early 1980s. They show scaleless glandular skin with a slightly bumpy texture, similar to that of hairless mammals or some amphibians. Since it occupied a point in the synapsid family tree close to where hair may have originated (somewhere in the Permian therapsids), it’s not clear if it was entirely hairless or if it had just secondarily lost some of it.
And Prolibytherium was probably the most striking of the lot.
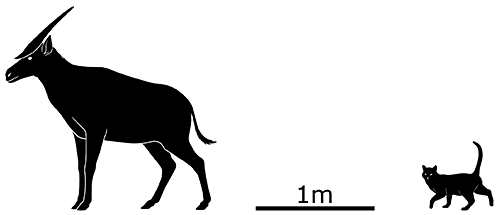
Two different species have been identified, with Prolibytherium magnieri here living in North Africa during the early-to-mid Miocene, about 17-16 million years ago. Its exact evolutionary relationships are uncertain but it was probably part of a group called climacoceratids, deer-like giraffoids which often had thorny branching ossicones that resembled antlers.
It stood around 1.2m tall at the shoulder (~4′), and exhibited dramatic sexual dimorphism – females had slender forked horn-like ossicones, while those of the males flared out into large wide flat shapes that resembled butterfly wings.
Heavy reinforcement in the bones of the back of the males’ skulls helped to support all the extra weight of those huge ossicones, and if they actually used the structures to fight with each other then this may have also provided some protection or shock absorption.
Modern ruminants are the only living mammals with bony headgear, with four different lineages each sporting a slightly different type: deer antlers, bovid horns, giraffid ossicones, and the prongs of pronghorns.
The protoceratids were an early group of North American ruminants whose relationships are uncertain, but may have been related to modern chevrotains. They were convergently deer-like in appearance, with teeth adapted for grazing on tough grasses – and along with having a pair of horns in the usual position on their heads, males also sported an additional pair of ossicone-like growths on their noses.
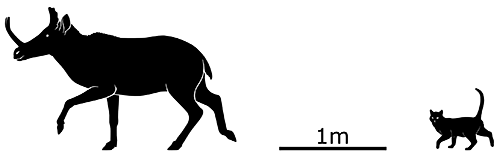
Synthetoceras tricornatus lived during the Late Miocene, around 10-5 million years ago, and was one of the largest protoceratids, standing about 1.1m tall at the shoulder (3’7″). Its two nose-horns were partially fused into a single long structure with a forked tip, which may have been used for sparring in a similar manner to the antlers of modern deer.
Synthetoceras tricornatus
Meanwhile on a different branch of the ruminant family tree, closer related to deer and giraffes, a group known as the palaeomerycids independently developed a similar sort of extra head appendage – but at the opposite end of their skulls.
These ruminants were a little more heavily built than the protoceratids, and specialized in feeding on soft vegetation in humid forest environments. They were a highly successful group, existing for almost 30 million years, ranging across Eurasia, Africa, and North America, and even ventured into South America during the early phases of the Great American Interchange.
Males had two giraffe-like ossicones above their eyes, along with a third crest-like one at the very back of their heads. In some species this formed a single central “horn” shape, while in others it forked out to each side. They also often had long saber-like canine teeth similar to modern water deer and musk deer, which were probably used for fighting while their elaborate headgear was purely for visual display.
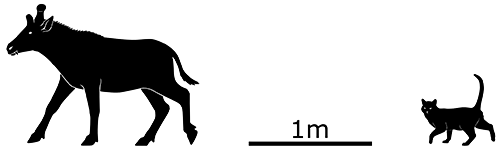
Xenokeryx amidalae lived in Spain during the mid Miocene, about 16 million years ago. It stood around 0.8-1m tall at the shoulder (2’7″-3’3″) and had a unique T-shaped “handlebar” crest which ended up inspiring its genus name – a reference to the similar shape of one of Queen Amidala’s headpieces in Star Wars, which was itself based on Mongolian imperial fashion.
Living in Nova Scotia during the Late Triassic, around 235-221 million years ago, Teraterpeton (meaning “wonderful creeping thing”) was first named in the early 2000s based on a skull and partial skeleton, with some additional skeletal material being described recently in 2019.
Its head had a confusing mix of anatomical features, with a long beak-like toothless snout at the front of its jaws, small sharp interlocking cheek teeth further back, a huge nasal opening, and a closed-up fenestra at the back of its skull making it look more like the skulls of marine reptiles.
It also had a lizard-like body, perhaps up to 1.8m long (~6′), with rather long slender limbs and large blade-like claws, and more anatomical weirdness in the pelvic region convergently resembling those of distantly related groups like rhynchosaurs and tanystropheids. It had a sprawling posture, but its hind limb musculature suggests it might have been capable of getting up into a more erect stance when walking, somewhat similar to modern crocodilians’ “high walk” gait.
It was clearly quite an ecologically specialized animal, but quite what it was specialized for is still uncertain. It was presumably a herbivore like its close relatives, but it must have been eating a very different diet with its long beak, and its deep claws could have been used for scratch digging to get at roots and tubers.
Another possibility it that it could have been an insectivore with a diet similar to modern aardvarks or armadillos, probing with its beak and digging with its claws for insects, grubs, and other invertebrates. Since termite-like social insect nests do seem to have existed around the same time, it might even have been one the earliest known animals to specialize in myrmecophagy.
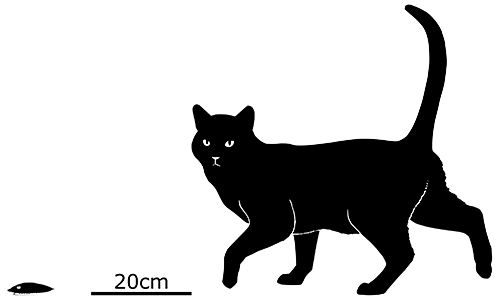
Trilobites were one of the most successful groups of early animals, existing for over 300 million years – and during that time they developed a huge diversity of weird heads, with various arrangements of spines, horns, eyestalks, and even long snouts and tridents.
But perhaps one of the oddest was the genus Odontocephalus, known mainly from the early-to-mid Devonian and represented here by Odontocephalus aegeria.
Living about 390 million years ago in northeast North America, this trilobite grew up to around 9cm long (3.5″). And although it wasn’t overall very elaborately ornamented, the front margin of its head had a row of extensions that flared out to meet at their tips, forming something resembling the cowcatchers used on trains.
The actual function of this structure is unknown. It might have been purely used for visual display since trilobites had excellent vision – but Odontocephalus was also a fast-moving bottom-dweller, and its “cowcatcher” may have served the same sort of purpose as its modern equivalent, deflecting small obstacles in its path as it trundled along the seabed.















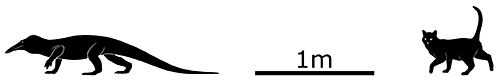
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}