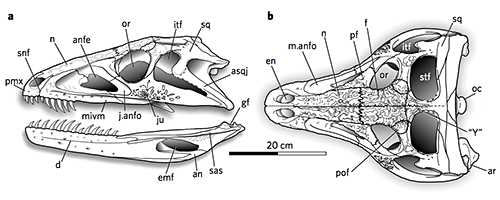
Medusaceratops lokii, a ceratopsid from the Late Cretaceous of Montana, USA (~77.5 mya).
About 6m long (19′8″), it had long brow horns and large curved spikes on its frill – an arrangement very similar in appearance to the centrosaur Albertaceratops, and initially its fossils were misidentified as belonging to that particular ceratopsid. But in 2010 it was recognized as a different genus, and based on some partial frill remains it was classified as a very early chasmosaur (a different branch of the ceratopsids which includes Triceratops), related to other early forms like Mercuriceratops.
Its genus name was based on the snake-haired Medusa from Greek mythology, while its species name comes from the Norse trickster god Loki – both in reference to the years of confusion about the identity of Medusaceratops’ fossils, and the distinctive curved horns on the helmet of Marvel’s Loki.
And, true to its name, the confusion wasn’t over yet.
Recently more fossil material and a new study have shown it was still being misclassified. Now it seems like Medusaceratops was actually part of the centrosaur lineage all along, and was indeed a very close relative of Albertaceratops.
It also turns out that what were thought to be numerous Albertaceratops fossils found in the same location were all just even more Medusaceratops. Instead of a mixture of two different ceratopsids there’s a single big bonebed representing some sort of mass-mortality event of only this one animal.
Similar mass bonebeds have been found for other centrosaurs in the same area and around the same age. Perhaps there were frequent flash floods at the time, or they were attempting to migrate across fast-flowing rivers like some modern animals, but we still don’t actually know for certain why they died en masse so frequently.











{kind=link}