It’s time for a little bit more recent PBS Eons work this week:


• The metatherian predator Arctodictis and the litoptern ungulate Thoatherium from “The Mystery of South America’s False Horses”
Paleontology and science illustration

It’s time for a little bit more recent PBS Eons work this week:
• The metatherian predator Arctodictis and the litoptern ungulate Thoatherium from “The Mystery of South America’s False Horses”
Albireonids were an early branch of the delphinoid whales, with their closest living relatives being modern oceanic dolphins, narwhals and belugas, and porpoises. Known from temperate latitudes of the North Pacific Ocean between the late Miocene and the late Pliocene, about 9-2.5 million years ago, their fossil remains are very rare in coastal deposits and they seem to have primarily been offshore open ocean animals.
Albireo whistleri is the best known member of this family, represented by a near-complete skeleton from what is now Isla de Cedros in Baja California, Mexico, dating to the late Miocene between about 8 and 6 million years ago. It was a rather small dolphin, around 2.5m long (~8’2″), with a stocky body, fairly broad flippers, and skull anatomy with some convergent similarities with the modern Dall’s porpoise.
Interestingly these dolphins also seem to have frequently had pathological neck vertebrae, with both Albireo whisteri and the younger species Albireo savagei from California, USA, showing unusually asymmetrical atlas bones – but on opposite sides to each other. This might be due to illness or injury earlier in life, or possibly be evidence of some sort of “handedness” with individuals preferring to perform some actions more with one side of their body than the other.
Continue reading “Albireo”Romaleodelphis pollerspoecki was a dolphin-like toothed whale related to the ancestors of both modern oceanic dolphins and beaked whales, living in coastal waters covering what is now Austria during the early Miocene about 22 million years ago.
Although only known from a single fossil skull, this cetacean was probably around 3m long (~9’10”). It had a long snout lined with over 100 small pointed uniformly-shaped teeth, and the bony walls of its inner ears were well-preserved enough to show that it was able to hear narrow-band high frequency sounds – a specific form of echolocation that has convergently evolved multiple times in various modern and extinct toothed whale lineages.
Based on the presence of ancient river-mouth deposits in the area where Romaleodelphis was found, it may potentially have been capable of traversing between marine, brackish, and freshwater environments similar to the modern franciscana.
Continue reading “Romaleodelphis”Okay, let’s finish up this year’s round of speculative evolution concepts with one last post before we get back to normal paleoart content.
(…what do you mean it’s November 3rd? how did that happen?)
Belial Lyka asked for a “marine proboscis monkey with a somewhat buoyant nose”:

A descendant of modern proboscis monkeys, Phusarhinus beliallykae is a large semiaquatic primate found in shallow marshy coastal areas of what was once Southeast Asia. Around 4m long (~13′) it has a lifestyle somewhat similar to ancient early sirenians, feeding on soft aquatic plants and hauling out onto land to rest.
Its dense rib bones and long flipper-like grasping forelimbs make it rather front-heavy, allowing it to naturally float with its head and arms hanging down closer to the bottom for energy-efficient foraging. When it needs to resurface to breathe it shunts air from its lungs into its large inflatable nasal sacs, altering its buoyancy enough to tip its head back up towards the surface.
Unlike its ancestors the elaborate nasal structures are found on both males and females – although they’re more brightly colored in males and are also used for visual courtship displays and as resonating chambers for loud booming calls.
And somebody who only gave their name as “bunny” suggested a “parasitic rodent”:

Sanguichelonamys bunnyi is a highly unusual descendant of a rakali-like semiaquatic rodent that had a symbiotic relationship with early members of the Phusarhinus lineage. The rodents initially just removed algae and external parasites from the bodies of the increasingly bulky aquatic monkeys, but things have recently started to turn more parasitic.
At just 3cm long (~1.2″) Sanguichelonamys is one of the smallest mammals to ever exist, with a wide flattened body and sharp hooked claws used to cling onto its host monkey’s thick skin. Although it still does remove other parasites, during haul-out periods it will also use its sharp incisors to deliberately enlarge the wounds left behind – or even open up new ones – and directly feed on fresh blood from its host.
The thickened keratinous skin along its head and back has a specialized hydrophobic surface that traps a layer of air while underwater, acting as a “rebreather” bubble similar to that of water anoles. Along with the ability to drastically slow down its metabolism and respiration rate, this allows Sanguichelonamys to survive being submerged during its host’s lengthy foraging dives.
@poshtearexdoodles asked for “African wild dogs evolved to fill the larger pack animal niche of lions”:
Although in this particular timeline the endangered and fragmented wild painted dogs in Africa didn’t survive far past modern times, that wasn’t the final extinction of the species – captive groups persisted, and after the [REDACTED] of humans an escaped population managed to carve out a niche for themselves in the grasslands of southern North America.

Their most notable descendant is Megainolycos poshtearexi. At nearly 1m tall at the shoulder (~3’3″) it’s one of the largest canids to ever exist, comparable to present-day lions in size and general ecology, with short thick bone-crushing jaws convergent with those of hyenas.
It also retains the complex variable coat coloration of its ancestors, with each individual having a unique disruptive pattern of black, white, and brown blotches.
Much like its ancestors it’s a highly social pack predator specialized for endurance hunting. Along with some surprisingly horse-like limb ligament morphology, its feet now sport hoof-like nails rather than claws, and it pursues prey over long distances to the point of exhaustion – mainly targeting the larger ungulates across its prairie habitat such as hogs, horses, bovids, and deer.
Roy (@roygattero) requested “ratel becoming the new african big predator”:

Perforictis royi is the latest in the long and venerable tradition of various synapsid lineages discovering the ecological niche of “big stabby saber-teeth“.
A descendant of the modern ratel/honey badger, it stands around 80cm tall at the shoulder (~2’8″), and convergently resembles the feline-like build of some of its more ancient relatives. Inhabiting the tropical forests of the rifted-off island continent of East Africa, it’s an ambush predator specializing in tackling larger prey – primarily ungulates and primates, but also occasionally giant rodents and hyraxes.
Usually cooperatively hunting in mated pairs, these mustelids stalk close to their targets before attacking, with one individual focusing on toppling and immobilizing their target while the other positions itself to deliver a swift precision killing bite to the throat with its saber-teeth and powerful neck musculature.
Perforictis scent marks its territory using extremely pungent secretions from its anal pouch rubbed onto vegatation, along with making loud scream-like vocalizations.
GayCoonie suggested a “future legless mustelid”:
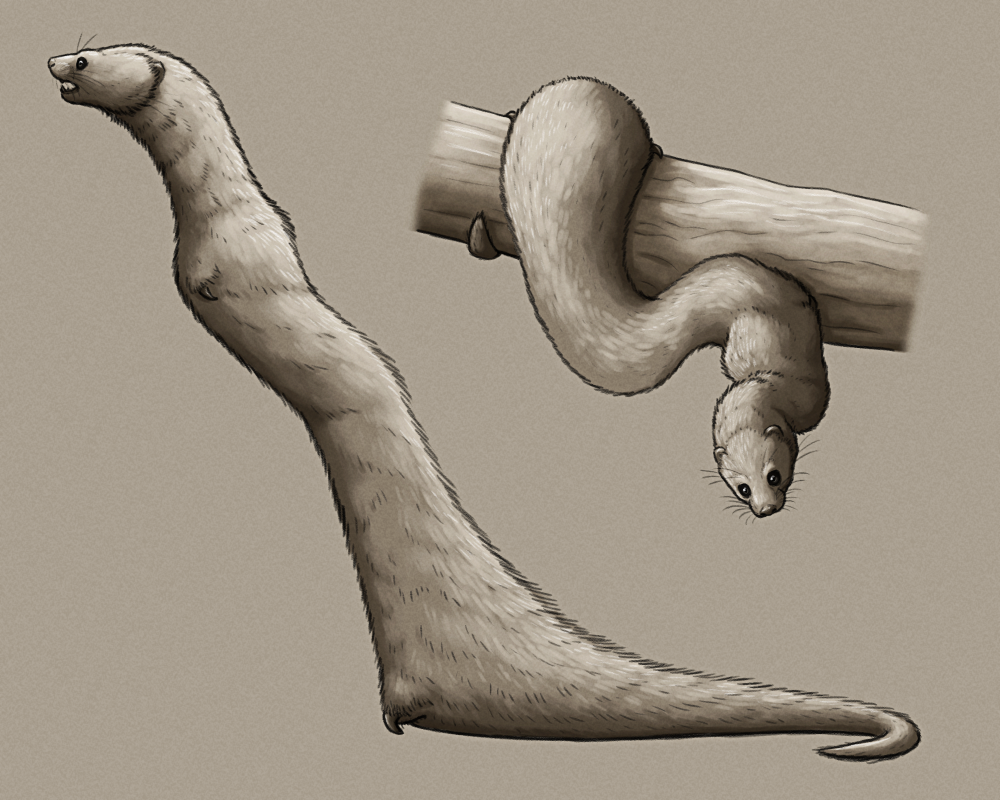
Descended from an arboreal offshoot lineage of the modern least weasel, Maximagale gaycooniei is a bizarre mustelid that appears to have converged on the lifestyle of ambush-hunting tree snakes, evolving in northern latitudes where actual snakes are largely absent.
Growing to about 1m long (3’3″), it’s not truly legless but its limbs are all reduced down to tiny vestigial single-clawed spurs, which are used to help anchor its body while climbing and as claspers during mating. It moves around with a distinctive inchworm-like looping gait, alternating grasping and releasing with its front and hind spurs.
Its build is bottom-heavy, with most of its mass concentrated in its thicker back end, and its tail is semi-prehensile. It clings to trees with its body bunched up, camouflaged with cryptic coloration, and rapidly whips its long flexible front half out to snap its powerfully-muscled jaws at prey – such as insects, birds, lizards, frogs, small mammals, and pretty much anything else that comes within its reach.
It will also opportunistically raid the nest of birds and arboreal mammals.
Due to the less frequent meals its ambush-hunting tactics provide, it has a much slower metabolism than its ancestors, and it conserves energy with daily periods of torpor and longer hibernation during the colder months of winter. It has also retained its ancestor’s tendency to seasonally shrink its brain size to reduce energy requirements even more.
Someone who identified themself only as Pendrew asked for a “ruminant-like Old World Monkey”:

After much of East Africa rifted off into a separate continent, shifting climate turned the alpine grasslands of what was once the Ethiopian Highlands into into warmer subtropical forests – and the highly terrestrial grass-eating geladas that inhabited the region adapted to new sources of food.
Yedenigelada pendrewsii is a large quadrupedal herbivorous monkey, about 1.5m tall at the shoulder (~5′). It has a specialized pseudoruminant digestive system with a three-chambered stomach, similar to that of camelids, and it occupies an ecological niche convergent with the ancient chalicotheres, selectively browsing on trees and shrubs while sitting upright and using its long clawed forelimbs to pull branches within reach.
Unlike its highly social ancestors this species is mostly solitary, although during the breeding season groups of males come together in leks to compete for female attention. Displays consist of inflating large colorful throat pouches to make loud resonating calls, and flipping upper lips to bare teeth and gums.

An anonymous submitter asked for a “buoyant ungulate that runs atop the sea”:
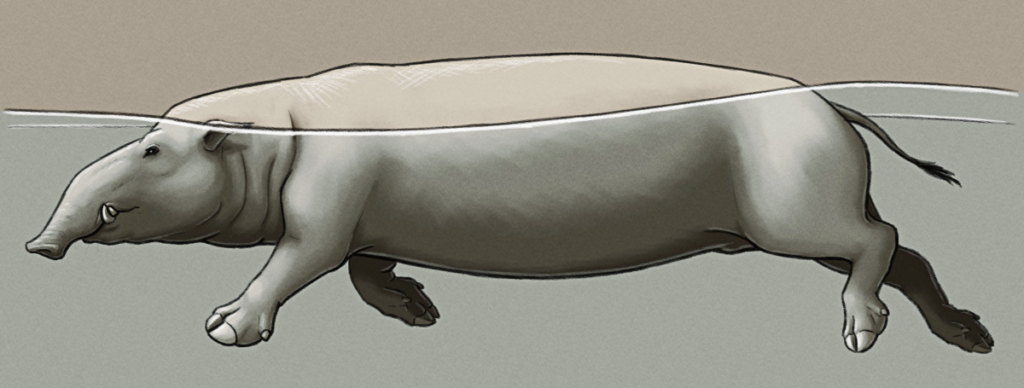
Pontoporcus plotus is a 1.5m long (~5′) amphibious pig descended from a feral population of domestic pigs left on a small tropical island. After inadvertently wreaking havoc on much of the local ecosystem, its ancestors eventually turned to a more marine-based lifestyle foraging along beaches and in coastal waters.
Naturally highly buoyant, Pontoporcus actually floats so well that it’s mostly limited to the water’s surface, unable to dive to any significant degree. But despite this it’s a fairly good swimmer, using broad hooves with wide fleshy pads to paddle itself along in an aquatic trotting- or running-like gait.
It forages both on land and in the water, mainly eating soft vegetation and marine plants, but much like its ancestors it will also opportunistically feed on whatever smaller animals it can catch or scavenge. Its semi-prehensile trunk-like snout is used to grasp at food items, to probe and root around in soft sediment, and as a snorkel.
Its hairless skin is very susceptible to sunburn, but it secretes a thick oily red-brown substance (similar to modern hippo “blood sweat”) that acts as a natural protective sunscreen.
These pigs are accomplished island-hoppers, regularly traversing the relatively shallow seas all along their island chain – but their natural flotation and long fat bodies also make them prime targets for large aquatic predators attacking from below, so these journeys tend to involve groups of Pontoporcus “running” along the sea surface aiming for their next destination as fast as they possibly can.
It’s September, it’s #Spectember, and I’m still plugging away at that big ol’ pile of speculative evolution idea submissions from a few years ago.
I will never be free.
Much like last year I’m not setting a definite posting schedule for this month; it’ll just be whenever and whatever I can manage to get done.
(Also, a reminder: I’m still not currently taking new requests!)
So let’s get started with an anonymous submission that requested a “swimming piscivorous canid”:

Descended from the short-eared dog (a species that in modern times already has partially-webbed paws and eats a large proportion of fish in its diet), Pelagicyon salsus is a 2m long (~6’6″) semi-aquatic piscivorous canid with a stocky body, short webbed limbs, and a long flattened tail.
The back of its skull is very wide, anchoring its thick neck musculature and accommodating huge cheekbones with powerful jaw muscles, but in contrast its snout is elongated and slender – a combination of features that allows it to sweep its toothy jaws through the water to rapidly snap at fish in a similar manner to gharials.
Most other members of its lineage inhabit freshwater rivers and swamps, but Pelagicyon is an unusual marine offshoot that has developed enough salt tolerance to swim, feed, and even drink exclusively in seawater.

