

The rhino-like toxodontids from earlier in this series weren’t the only weird-headed South American ungulates. Another group known as the litopterns evolved in a different direction, becoming long-legged fast-moving animals convergently filling the same sort of ecological niches as modern horses, deer, bovids, camelids, and giraffids.
Macrauchenia patachonica was one of the strangest members of this lineage, living from the Late Miocene to the end of the Pleistocene, between about 7 million years ago and just 12,000 years ago.
It stood around 1.8m at the shoulder (5’11”) and resembled a large camel or llama with thee-toed hoofed feet, but its head was… confusing.
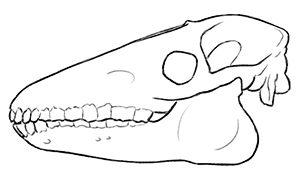
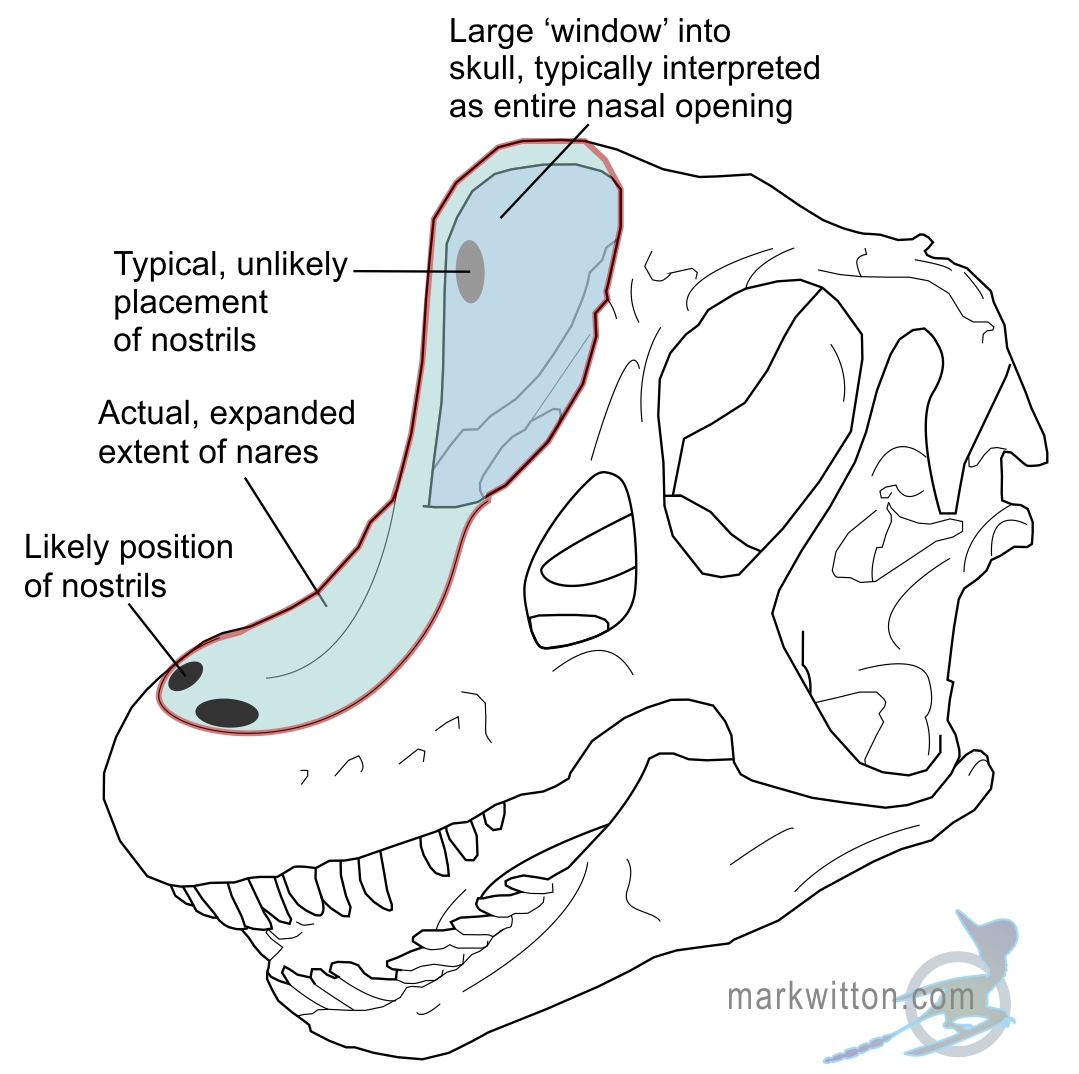
Its skull had a bizarre combination of features, with a shape closer to a sauropod dinosaur than a mammal, a cartoonish-looking set of teeth, and its nostrils set up high above its eyes, more like a cetacean blowhole than a terrestrial herbivore.
Due to its retracted nostrils it’s commonly been restored with an elephant-like or tapir-like trunk. And while a trunk gives Marauchenia a wonderfully weird and memorable appearance, there’s just one problem with that interpretation.
There’s no evidence for it.
Aside from its nostrils being far back on its head, it didn’t have any other features associated with anchoring the complex musculature of a trunk. In fact, a recent study found that its skull characteristics were much closer to those of moose than tapirs!
It seems more likely that it had a moose-like bulbous fleshy nose – possibly giving it an enhanced sense of smell or functioning as a resonating chamber – perhaps with slightly retracted external nostrils like a giraffe or sauropod to prevent it from being stabbed in the nose when feeding on spiky vegetation.
Whatever it was doing with its weird schnoz, it was clearly a highly successful species, since it was found across most of South America in a wide range of habitats.













_2_(15220449947).jpg){kind=link}
{kind=link}
.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}