

Soft-bodied annelid worms only very rarely fossilize, so the group’s origins during the Cambrian Period are still rather poorly understood. So far about thirteen different species have been found in sites of exceptional preservation, showing that even very early on in their evolution these worms had already diversified into a wide range of ecologies including bottom-feeders, carnivores, swimmers, tube-builders, and even symbiotes sharing living space with early acorn worms.
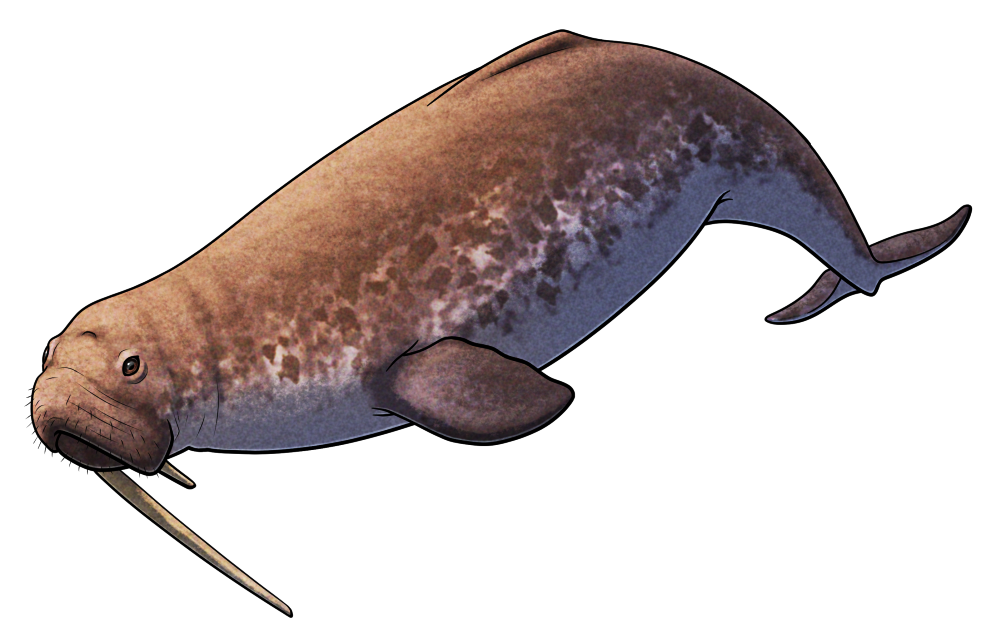
Ursactis comosa here adds a fourteenth species to the list. Found in a newly-discovered outcrop of the 508-million-year-old Burgess Shale fossil deposits in western Canada, it’s known from nearly 600 specimens clustered together in several large groups, making it the current best-known and most numerous of all Cambrian annelids.
Up to about 1.5cm long (~0.6″), it was a polychaete-like worm bearing bundles of long bristles. There was a pair of large sensory palps on its head, and its body was made up of an unusually small number of segments – just 10, with larger individuals just increasing the size of their segments instead of adding on additional ones like most modern annelids.
Unlike other Cambrian annelids it also shows some evidence of basic tagmatization, differentiating some of the rear segments of its body with much longer bristles.
The large numbers of Ursactis found preserved in one place suggests these worms were exhibiting some sort of swarming behavior. Since ages from juveniles to fully-grown adults are represented together, and their anatomy indicates they were crawling detritivores, they were probably all taking advantage of a particularly nutrient-rich patch of seafloor at the time they were abruptly buried in a mudslide.















{kind=link}
{kind=link}
{kind=link}