Horseshoe crabs are famous examples of “living fossils“, having changed their external appearance very little over hundreds of millions of years. But some fossil species were much more varied in shape than their morphologically conservative modern relatives, such as Austrolimulus fletcheri here.
Living in freshwater environments in what is now New South Wales, Australia, during the Middle Triassic (~247-242 million years ago), Austrolimulus had incredibly long spines on each side of its head, reaching a span of around 18cm (7″) – wider than its total body length!
The function of these spines is unclear, but they may have acted like a hydrofoil in fast-moving currents, or they may have served a defensive purpose by making Austrolimulus‘ carapace too wide and unwieldy for some predators to deal with.
Consisting of a partial jawbone and a humerus, and dating to the mid-Eocene (~47 million years ago), the remains clearly belonged to an early even-toed ungulate – but one much bigger than the rabbit-sized herbivores known from that time. This was something closer in size and build to a large modern pig, standing at least 1m tall at the shoulder (3’3″).
It was one of the earliest known large-bodied members of the group, and shows that these animals must have increased in size very rapidly during their early evolution, going from rabbit-sized to pig-sized within just a couple of million years.
The modern tuatara is the only living representative of an entire major lineage of reptiles known as sphenodontians – an evolutionary “cousin” group to all lizards and snakes, last sharing a common ancestor with them over 240 million years ago.
And during the Triassic and Jurassic these lizard-like animals were a widespread and diverse bunch, found worldwide and occupying many of the ecological roles that were later taken over by true lizards. They ranged from tiny insectivores to omnivores, relatively large herbivores, and specialized shell-crushers – and some even adapted to a fully aquatic fish-eating lifestyle.
Pleurosaurus ginsburgi here lived during the Late Jurassic, about 150-145 million years ago, in the warm shallow seas and lagoons that covered most of Europe at that time. Fossils of this particular species are known from southern France, with the closely related Pleurosaurus goldfussi found in both the same region and the German Solnhofen Limestone.
These swimming sphenodontians could grow to around 1.5 in length (~5′), with elongated bodies, pointed triangular snouts with retracted nostrils, short flipper-like forelimbs, and especially long eel-like tails. Soft tissue impressions also show scaly skin covering their bodies and a “frill” running along the top of the tail.
And one of the groups that rose to prominence during this time were the rhynchosaurs. Part of the archosauromorph branch of reptiles, they were closely related to the ancestors of crocodilians, pterosaurs, and dinosaurs, and evolved from small superficially lizard-like forms living in southern Africa during the very start of the Triassic, around 250 million years ago. But within just a few million years they became larger and bulkier, specialized for herbivory and scratch digging, and they soon spread all over Pangaea and became incredibly abundant in some fossil deposits.
Stenaulorhynchus stockleyi was one of larger member of this lineage, around 1.2m long (4’), known from Tanzania about 247-242 million years ago. It had a typical triangular rhynchosaurian skull, with wide deep cheeks supporting powerful jaw muscles and multiple rows of grinding teeth, along with a narrow hooked “beak” formed from the premaxillary bones of its snout.
Discovered in the late 1820s by pioneering paleontologist Mary Anning, the odd-looking fossil of the cartilaginous fish Squaloraja polyspondyla seemed to have characteristics of both sharks and rays.
It was initially thought to be a “missing link” transitional form between those two groups, but later it was identified as being something else entirely – it was actually part of the chimaera lineage, much closer related to modern ratfish, and its ray-like features were due to convergent evolution for a bottom-feeding lifestyle.
Living during the early Jurassic period, about 200-195 million years ago, Squaloraja fossils are now known from the south coast of England, southern Belguim, and northern Italy. Around 30cm long (1’), this weird fish had a massive wide flat snout that looked like an even more extreme version of the long noses seen in some of its modern relatives, and this enormous snoot would have been absolutely packed with sensory receptors to help it locate small aquatic prey hidden in the muddy seafloor.
Some specimens also have a distinctive long horn-like spine on their foreheads, and since these individuals also have claspers it seems like this was a sexually dimorphic feature. Much like the smaller head claspers on modern chimaeras, male Squaloraja probably used this “horn” to hang onto females’ pectoral fins during mating – and with it being such a large elaborate structure it may also have been used for visual display purposes, too.
A group of early tetrapods known as diadectids were some of the first land animals to become specialized herbivores, developing adaptations for the bulk processing of tough fibrous vegetation. They had powerful jaws, chisel-like front teeth, and grinding cheek teeth, and they grew to relatively large sizes for their time with bulky bodies supporting voluminous plant-fermenting guts.
Although usually considered to be reptilomorphs – “amphibian-grade” animals more closely related to amniotes than to modern amphibians – some studies have instead placed these early plant-eaters as being true amniotes related to the synapsids. Fossil trackways show they may have had amniote-like claws on their feet, and that their highly flexible lizard-like ankle joints allowed them to walk much more efficiently than other early tetrapods, possibly using a semi-upright gait, but these may be convergently evolved features. Since we don’t know whether they laid amniote-like eggs or if they instead spawned amphibian-style in water, it’s currently hard to tell for certain just what they really were.
Diasparactus zenos (sometimes alternately known as Diadectes zenos) was a diadectid that lived during the early Permian in New Mexico, USA, about 296 million years ago. Around 1.3m long (4’3”), it was only about half the size of its largest relatives, but it’s notable for having unusually high neural spines on its vertebrae – not quite long enough to be considered a sail, but more of a “high back” that may have supported powerful musculature or fatty deposits.
At the time this region was a river plain with a tropical climate, experiencing seasonal floods that turned the whole area into what’s known as “viesses” (a name based on the abbreviation “V.S.S.” standing for “very shallow sea”), vast shallow lake-seas that persisted for weeks or months at a time.
So this little animal has been interpreted as being semi-aquatic, swimming around and feeding on aquatic invertebrates and tiny fish and amphibians. Its skull had numerous pits around the front of its face, suggesting that it had a highly sensitive snout – probably whiskery, allowing it to hunt entirely by touch in dark murky water, but it’s also been proposed to have possibly had an electroreceptive sense similar to modern platypuses.
Brontornis burmeisteri was one of the largest flightless birds known to have ever existed, standing around 2.8m tall (9’2″) and estimated to have weighed 400kg (~880lbs).
But Brontornis might not actually have been a terror bird at all – it may have instead been a giant cousin of ducks and geese.
The known fossil material is fragmentary enough that it’s still hard to tell for certain, but there’s some evidence that links it to the gastornithiformes, a group of huge herbivorous birds related to modern waterfowl.
If it was a gastornithiform, that would mean it represents a previously completely unknown lineage of South American giant flightless galloanserans. And, along with the gastornithids and the mihirungs, it would represent a third time that group of birds convergently evolved this sort of body plan and ecological role on entirely different continents during the Cenozoic.
Non-tetrapod vertebrates like fish have spines that are much less differentiated, with just body and tail segments. So for a long time multiple distinct spine regions were thought to be something completely unique to tetrapods – a specialization developed early in their evolutionary history that served to better support their weight when moving around on land.
But one little fossil fish makes this idea… problematic.
Tarrasius problematicus lived during the early Carboniferous, about 345 million years ago, in shallow tropical marine waters in what is now southern Scotland. Around 9cm long (3.5″), it was an early type of ray-finned fish with a scaleless body and a long scaled eel-like tail with a single continuous dorsal fin.
Its spine shows five different regions all corresponding to those seen in tetrapods, despite it not being closely related to them. But unlike early tetrapods Tarrasius was no land-walker, with its lack of hind fins indicating it was instead a streamlined fully aquatic fast swimmer.
It’s not clear why this fish developed such an incredibly convergent backbone, but it may have helped to stiffen its body so its more flexible tail could provide more efficient thrust, swimming like a modern tadpole.
It also suggests that a pre-existing genetic basis for regionalization – specific patterns of Hox gene expression – was actually an ancestral trait for all bony fish or jawed vertebrates. Tarrasius and early tetrapods may have just happened to specialize their spines in the same way for different purposes, with only the tetrapods going on to see long-term evolutionary success with it.
They were previously known mainly from isolated teeth and jaw fragments, with some rare full skull material, but Adalatherium is remarkable for being represented by a complete skeleton.
And it’s turned out to be far stranger than anyone expected.
Living in northwestern Madagascar during the Late Cretaceous, about 70-66 million years ago, Adalatherium was one of the larger known Mesozoic mammals at around 60cm long (2′) – although the one known specimen seems to have been a juvenile, so mature individuals were probably slightly larger.
It was probably a marmot-like digging animal, excavating burrows with its large claws and powerful limbs, and since it likely evolved from ancestors that had become isolated on Madagascar over 20 million years earlier it had developed a very unusual mixture of both “primitive” and highly specialized anatomical features. It had more back vertebrae than any other known Mesozoic mammal, upright forelimbs, sprawling hind legs with bowed-out tibias, strong back and leg musculature, and a therian-like pelvis with epipubic bones.
And then there’s the snoot.
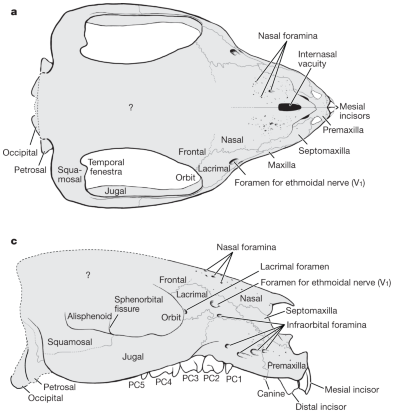
The snout region of Adalatherium‘s skull was pockmarked with a large number of foramina, holes that allow the passage of nerves and blood vessels through the bone. It had more of these than any other known mammal, and their presence suggests that it probably had a very sensitive upper lip and whiskery snout. Most mammals with a lot of whiskers just have one very big foramina, but Adalatherium seems to have evolved a different solution to the same problem.
It also had one other bizarre feature – a hole in the top of its nose. A large “internasal vacuity” between its nasal bones is a unique feature not known in any other mammal, and its function is a total mystery.
Since this hole was also surrounded by many foramina it may have supported some sort of soft-tissue sensory structure on top of its nose. So I’ve speculatively depicted it here with a leathery horn-like “shield”.
Adalatherium skull From fig 2 in Krause, D. W. et al (2020). Skeleton of a Cretaceous mammal from Madagascar reflects long-term insularity. Nature 581, 421–427. https://doi.org/10.1038/s41586-020-2234-8











{kind=link}