Toothed whales – the branch of cetaceans that includes modern dolphins, porpoises, beaked whales, and sperm whales – have surprisingly asymmetrical skulls, with some of the bones skewed to one side and just the left nostril forming their blowhole.
Some of the most obvious external manifestation of this lopsidedness can be seen in sperm whales, which have their blowhole at the front left side of their head, and in male narwhals, which usually have a single left-side tusk.
This sort of asymmetry first appeared in the skulls of early toothed whales around 30 million years ago. And since the highest amounts of wonkiness have gone on to develop in lineages that hunt in dark, cluttered, or murky waters, this suggests that the trait is somehow linked to the evolution of complex echolocation.
Some ancient members of the river dolphin lineage also had some additional unusual asymmetry, sometimes having slightly sideways-bending snouts.
Ensidelphis riveroi was one of the weirdest of these, living around the coasts of what is now Peru during the Miocene, about 19 million years ago. Around 3m long (~10′), it had a very long narrow toothy snout that curved distinctly off to the right along its length.
Expectation vs reality
It’s not clear what the function of this bend was, or even if the only known skull actually represents the normal condition for this species. But Ensidelphis’ bendy snoot might have been used to probe around in muddy seafloor sediment or to extract prey from crevices, possibly like an underwater version of the modern wrybill.
Antaecetus aithai was an early whale that lived during the late Eocene (~40 million years ago) in what is now Morocco, at a time when northern Africa was covered by a warm shallow sea.
It was part of the “basilosaurids“, some of the first fully aquatic cetaceans – traditionally considered to be a single defined group, but more recently found to be more of an “evolutionary grade” of multiple early whale lineages – and much like Basilosaurus it had elongated back vertebrae that would have given it a very long slender body shape.
Antaecetus also had a proportionally smaller head and smaller teeth than other basilosaurids, along with much denser bones and a stiffer spine that would have made it a rather slow swimmer with reduced maneuverability. It was also fairly small overall compared to most of its relatives, probably around 6m long (~20′).
It was probably a slow-moving coastal water animal somewhat like modern sirenians – except unlike manatees and dugongs it was carnivorous. Its relatively delicate teeth suggest it was feeding on soft-bodied prey like cephalodpods, and with its lack of speed it must have been some sort of ambush predator, waiting around for potential prey to come within striking range.
Towards the end of the Cretaceous, about 69 million years ago, the most diverse and numerous mammals in the northern hemisphere were the metatherians, close relatives of modern marsupials.
About the size of a modern mouse, around 10-15cm long (4-6″), and with teeth that suggest it was a shrew-like insectivore, this little metatherian lived in northern Alaska in what’s known as the Paaŋaqtat Province – a region with a distinctive population of endemic polar animals. At the time this area was located at an even higher latitude than it is today, around 80-85ºN, but due to a greenhouse climate it was also warmer, with no permanent ice and the average temperatures staying above freezing.
Unnuakomys was by far the most common mammal species in the Paaŋaqtat Province, represented by numerous fossil teeth and a few jaw fragments, and it also seems to have been the only metatherian living in the whole region. This may just be a preservation bias in the fossil record, but it might also indicate that Unnuakomys was uniquely specialized to endure the several months of continuous darkness each winter in its polar woodland environment, while other North American metatherians were restricted to more southerly latitudes.
And yet despite the huge diversity of gliding mammals, and their particular prevalence in tropical forests, there’s an entire continent famous for its rainforests that’s somehow completely lacking any modern examples: South America.
It’s not clear why the gliding lifestyle never took off in South America, but the continent is surprisingly devoid of any other gliding vertebrates, too. The only exceptions are a few species of flying frogs in the northwestern tropical forests around Colombia.
But back in the early Eocene, about 53-50 million years ago, there was at least one South American gliding mammal. Some fossil limb bones found in the Itaboraí Formation in southeastern Brazil look very much like those of a gliding mammal – long and thin, with a locking elbow joint, knees adapted for jumping, and flexible ankles typical of tree-climbers.
These remains haven’t been given a new scientific name, however, because there’s a good chance they belong to an already-described species. Fossils from Itaboraí are found disarticulated, broken, and with bones of multiple different species jumbled together, so most fossil mammals named from the site have been based on their more easily distinguishable teeth and jaw fragments.
The problem is matching those teeth with these bones.
Currently the best identity guess based on size is Gaylordia macrocynodonta. This mammal would have been around 30cm long (1′), about the size of a modern rat, and had distinctive large canine teeth. It used to be classified as a marsupial related to opossums, but more recent studies have found it to have actually been a marsupialiform metatherian instead, much more closely related to Pucadelphys and sparassodonts than to any modern true marsupials.
Gaylordia‘s crushing molars suggest it was carnivorous, able to crunch through bones or hard-shelled invertebrate prey. This would be a very unusual diet for a gliding mammal, since most other mammalian gliders are herbivores or omnivores – the only other known predatory examples were the volaticotheres over 110 million years earlier.
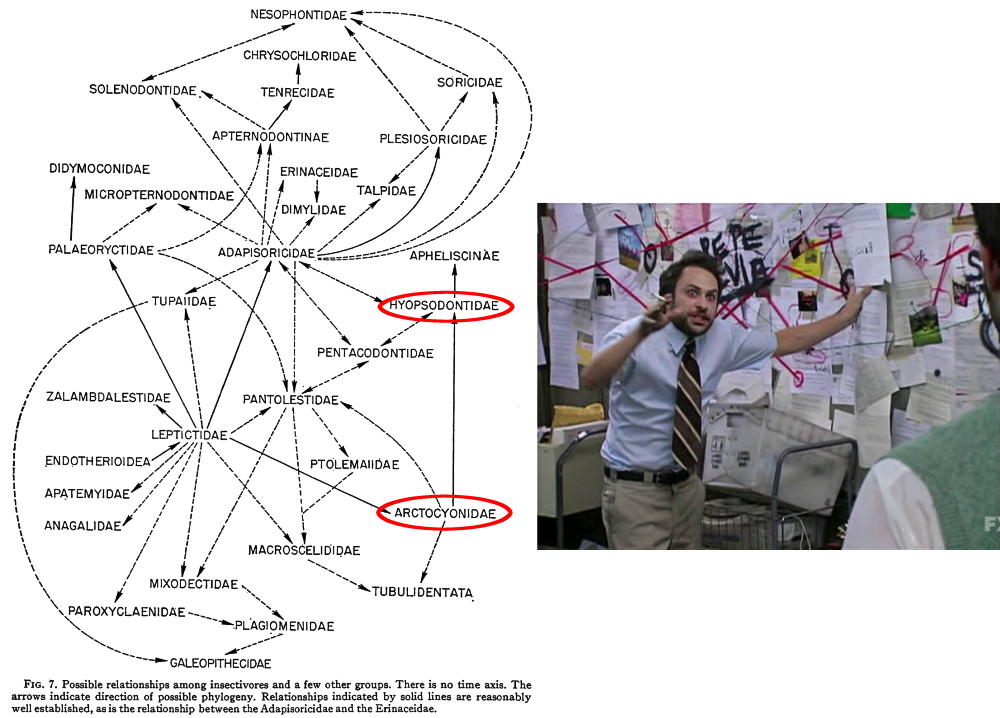
“Insectivora” was a wastebasket taxon so bad it had to be revised multiple times, but there’s another particularly infamous case in mammal taxonomy that’s still in the process of being resolved – the “condylarths“.
This group was first created in the early 1880s, during the Bone Wars, and initially was just a subgroup of odd-toed ungulates containing only the phenacodontids. But just a few years later Condylarthra was promoted up to its own order, and groups like the periptychids and hyopsodontids were added in too.
Then over the next few decades various groups were added and removed from the condylarths, most notably with the mesonychids and arctocyonids being brought in from their previous position with the creodonts.
By the mid-20th century the condylarths had become a big convenient dumping ground for any and all “primitive” ungulate-like mammals that didn’t easily fit into any modern groups, ranging in age from the early Paleocene through to the early Oligocene. But it soon became apparent that they had the same problem as the “insectivores” – there weren’t really any unique anatomical features that united all these animals together.
They generally had rounded-cusped molar teeth and hoof-like toes, but they also had rather generalized “primitive mammal” features and a diverse range of ecologies. Some were small herbivores, but others were coati-like or dog-like omnivores, and some were even bear-sized carnivores.
From left to right, top row: Hyopsodus lepidus (hyopsodontid), Meniscotherium chamense (phenacodontid), Arctocyon primaevus (“arctocyonid”). Bottom row: Ectoconus ditrigonus (periptychid), Mesonyx obtusidens (mesonychid)
It wasn’t even clear how the various different condylarth groups were actually related to each other. The best guess was that arctocyonids had arisen from within the “insectivores”, with a Protungulatum-like form as the common ancestor of all the other condylarths. Where exactly modern ungulates had then evolved from within the condylarths was also still uncertain.
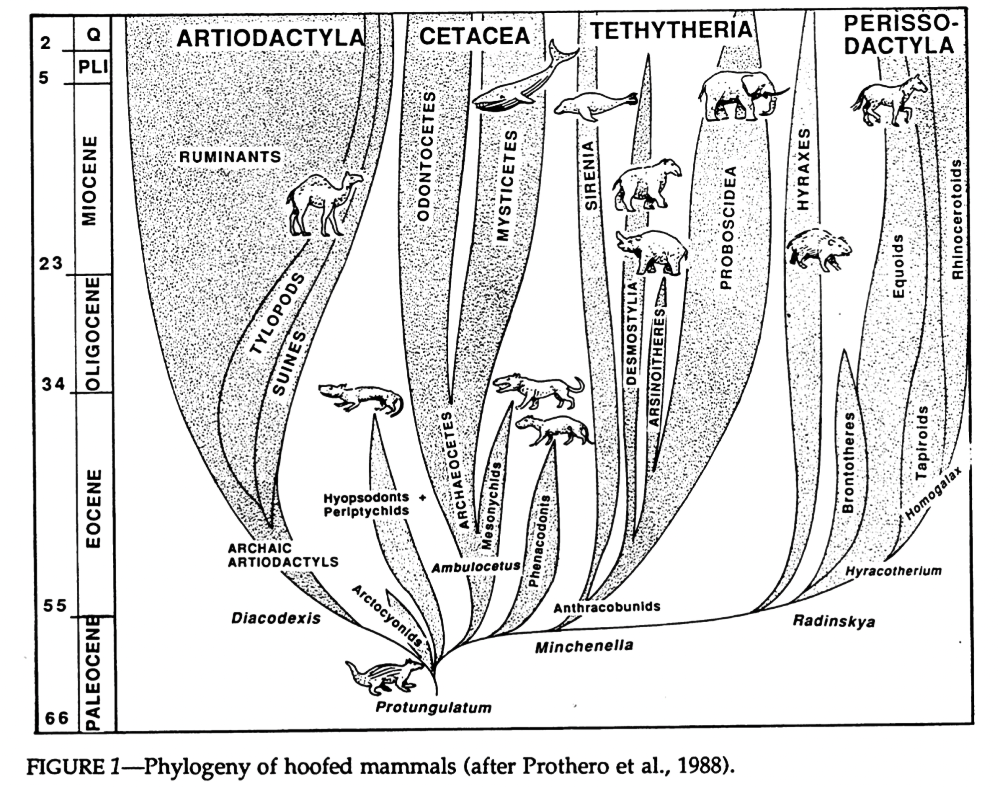
Cladistic analysis in the 1980s began to tackle the confusing pile of assorted condylarths, and showed that they weren’t the single ancestral source of all modern ungulates, but instead a loose collection of several unrelated groups from all over the ungulate evolutionary tree. Arctocyonids, periptychids, and hyopsodontids were placed as early “primitive” lineages, phenacodontids were loosely linked with the ancestors of odd-toed ungulates once again, and mesonychids were considered to be the ancestors of whales.
And, once again paralleling the mess of the “insectivores”, it wasn’t until genetic methods became available in the late 1990s that larger-scale ungulate relationships began to be properly resolved. The paenungulates (elephants, hyraxes, and sirenians), which had been traditionally considered to be a major branch of ungulates, were removed entirely and reclassified as afrotheres. And, along with some new fossil discoveries, whales were recognized as having actually evolved from within the even-toed ungulates instead of from mesonychids.
This shake-up threw the still-problematic “condylarth” classifications back into question – with some “condylarths” turning out toalso be afrotheres instead of true ungulates.
Today the actual relationships of the main “condylarth” ungulate families are still in the process of being figured out, and there’s a lot of remaining uncertainty and disagreement about them.
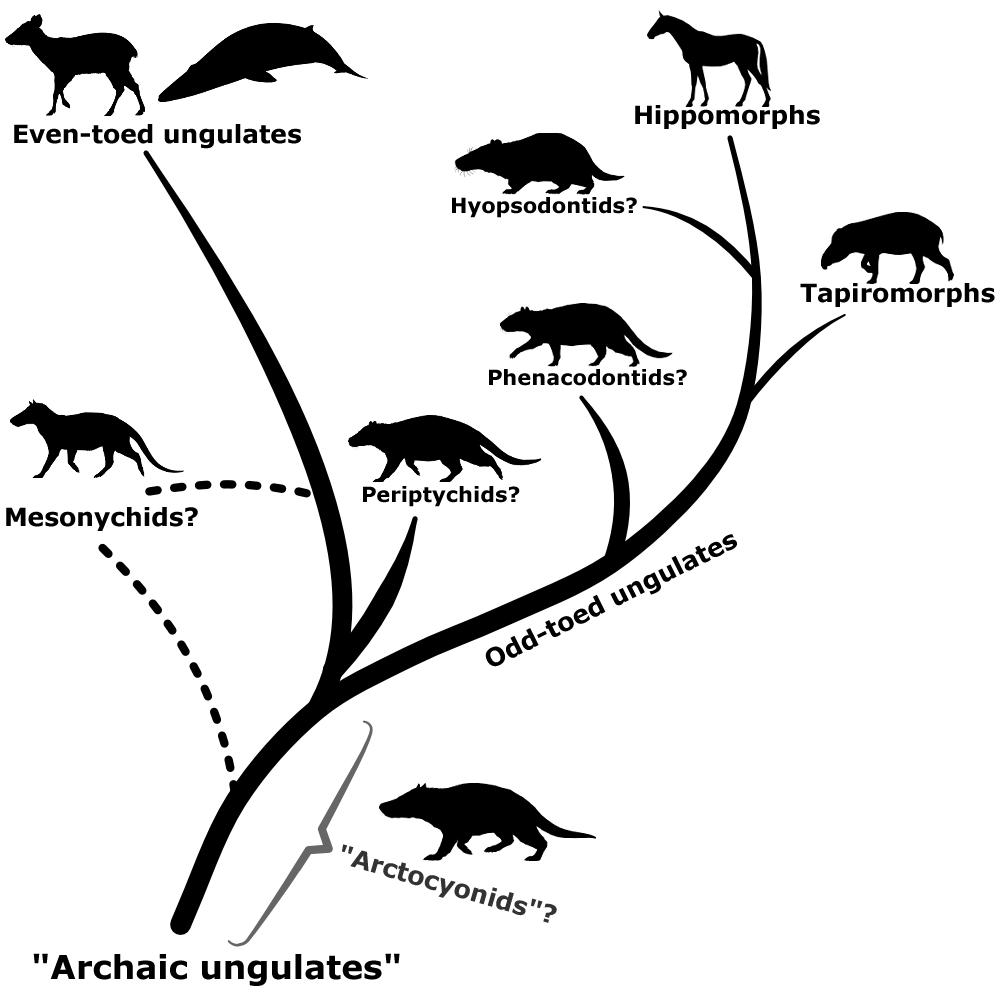
Phenacodontids seem to have mostly maintained their traditional position as early odd-toed ungulates, and hyopsodontids may potentially be part of this group too – possibly as members of the hippomorph lineage, closely related to horses and brontotheres. Arctocyonids might be a wastebasket themselves, with some studies finding them to be a mix of several different archaic ungulate lineages. Periptychids may have links to the even-toed ungulates. The mesonychids, meanwhile, are now generally considered to be a separate order from the traditional “condylarths”, and may be either an early branch of the even-toed ungulates or much more basal ungulates closely related to the “arctocyonids”.
Since the term “condylarth” no longer has any real taxonomic meaning some paleontologists have proposed replacing it with “archaic ungulate” to distance from the historical messiness of the old name. But this hasn’t really caught on, and many papers still use “condylarth” in a very loose sense to refer to an “evolutionary grade” of early ungulates of unclear evolutionary affinities.
And while that’s the last main entry for this month, we’re not quite done yet. There’s still one weekday left in October, and after digging through so many taxonomic garbage cans there’s only one place we can go now.
The rodent-like multituberculates were a major lineage of mammals that were only distantly related to modern marsupials and placentals. They originated around the time of the mid-Jurassic (~168 million years ago), survived through the end-Cretaceous mass extinction, and went on to become one of the most diverse and successful types of mammal in the Paleocene. After that point they began to decline, and after anw over-130-million-year-long run they went extinct* in the early Oligocene (~33 million years ago).
(* Except, possibly, in South America, where an enigmatic fossil known as Patagonia peregrina may represent a multi surviving as recently as about 18 million years ago in the early Miocene.)
First discovered in North America in the 1880s, Catopsalis foliatus was part of a group of multituberculates called taeniolabidoids. These multis got significantly larger than the rest of their kind – averaging beaver-sized but with some species getting up to at least capybara-sized – and were some of the first mammals to evolve into relatively big herbivores after the extinction of the non-avian dinosaurs.
Catopsalis foliatus
Catopsalis was named based on a partial jawbone and a few teeth, and over the next century or so various other similar-looking fossils from both North America and Asia were added into the genus as additional species. Eventually Catopsalis contained eight different species, ranging over about 10 million years from the late Cretaceous to the early Eocene – not especially big compared to some other wastebaskets we’ve looked at this month, but it was still a problem, muddying up attempts to understand the actual evolutionary relationships and biogeography of the taeniolabidoids.
Cladistic studies in the 1980s showed that Catopsalis was paraphyletic, made up of at least five separate lineages, and a few of them were subsequently renamed and reclassified. The Cretaceous Asian forms became Djadochtatherium and Catopsbaatar, and are now considered to be part of a different lineage of multis known as djadochtatherioids, while one of the remaining North American species then became Valenopsalis.
…But a couple of other new Catopsalis species have also been named in the meantime (one as recently as 2018), so there are still seven different species that need sorting out in this particular wastebasket.
Cetotheres were a group of small baleen whales, one of three major lineages of these cetaceans alongside the rorquals and the right whales. They first appeared in the fossil record in the mid-Miocene, about 14 million years ago (but are estimated to have actually originated 10-15 million years earlier), and disappeared during the Pleistocene about 2 million years ago.
First recognized in the mid-19th century, for a long time the cetotheres were used as a wastebasket for all fossil baleen whales that didn’t clearly fit into any modern whale families. By the start of the 21st century nearly 30 different genera representing numerous different species were all lumped into the group – and the genus Cetotherium was another wastebasket in itself with at least 12 assigned species, many of which were based on fragmentary or dubious remains.
Ciuciulea davidi
This was finally cleaned up in the 2000s, when a revision of the cetotheres cut the group down to just 6 genera. Since then a handful of additional new genera and species have been named, and while a few polyphyleticCetotherium species may still need tidying up the cetotheres have overall gone from being a total taxonomic mess to actually being one of the best studied groups of fossil baleen whales.
Their exact evolutionary relationships with each other are still in flux, but the most surprising discovery from the improved understanding of these ancient whales is that they might not be extinct after all.
At this point it seems fairly likely that the pygmy right whale really is either the last surviving representative of the cetothere lineage, or at least is a very close evolutionary “cousin” (a “cetotherioid”) closer related to them than to any other modern baleen whales.
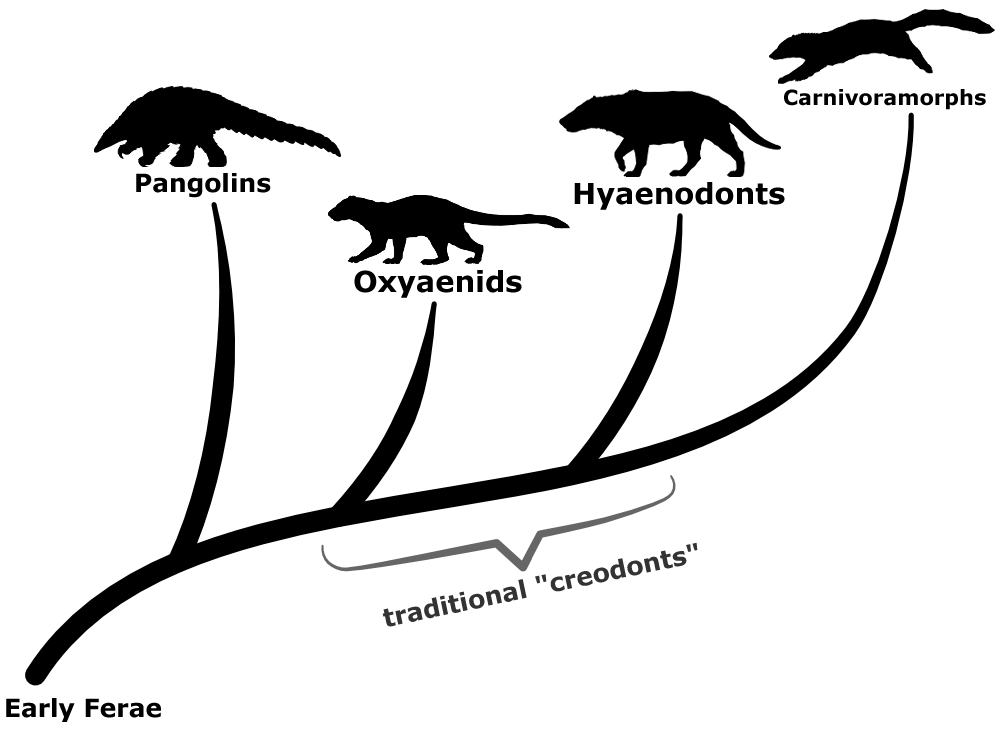
Creodonts were some of the earliest predatory placental mammals to evolve after the extinction of the non-avian dinosaurs, first appearing in the mid-Paleocene about 60 million years ago. Represented by two main lineages – the oxyaenids and the hyaenodonts – they ranged across North America, Eurasia, and Africa, and were the dominant large carnivorous mammals until the end of the Eocene (~34 million years ago), with forms like Sarkastodon being some of the biggest mammalian land predators of all time.
After that point they started to decline over most of their range, gradually being replaced by early carnivorans – but the hyaenodonts retained their dominance for a while longer in Africa, diversifying during the Oligocene and early Miocene and producing more giant apex predators. The last known representatives of these animals survived in Asia until the late Miocene, just 9 million years ago, ending an impressive run that had lasted for most of the Cenozoic.
This grouping was originally named in the 1870s to encompass just the oxyaenids and Didymictis (a genus now considered to be a viverravid). Just a few years later hyaenodonts, miacids, arctocyonids, leptictids, and mesonychids were all lumped in, too – and at one point creodonts were even a part of the massive insectivoran mess before instead being classified as ancestors of the carnivorans.
During the first half of the 20th century creodonts were recognized as actually being a loose collection of mostly-unrelated mammals, and over the next few decades various groups were gradually removed and reassigned to other parts of the mammal family tree. Towards the end of the century most of the creodont wastebasket had been cleared, and just the oxyaenids and the hyaenodonts were left as two branches of one seemingly distinct creodont lineage.
The cougar-sized oxyaenid Patriofelis ferox (left) & the bear-sized hyaenodont Hyaenodon gigas (right)
…But their evolutionary relationships were still a problem.
They’d been traditionally considered to be early carnivorans, but although they had flesh-slicing carnassials the creodonts’ versions of these teeth weren’t quite right. Different teeth in their jaws had been specialized for this function compared to those of true carnivorans – with oxyaenids and hyaenodonts having slightly different arrangements compared to each other, too – suggesting a lot of convergent evolution rather than shared ancestry.
By the 1990s it wasn’t clear anymore if the oxyaenids and hyaenodonts were even closely related to each other, or what type of mammal they actually were.
But over the last couple of decades the consensus seems to have become that creodonts weren’t a single natural group, but that they were still related to carnivorans – oxyaenids and hyaenodonts were actually two separate offshoots of the Ferae, forming an evolutionary grade of stem lineages between pangolins and the carnivoramorphs.
Most modern meat-eating placental mammals are carnivorans, a group that contains two distinct lineages: the feliforms (cats, hyenas, mongooses, viverrids, civets, linsangs, and euplerids) and the caniforms (dogs, bears, seals, raccoons, and mustelids).
The closest living relatives of these animals are pangolins, and their last common ancestor probably lived sometime between the Late Cretaceous and early Paleocene. But the actual early evolutionary history of the carnivorans themselves is rather murkier.
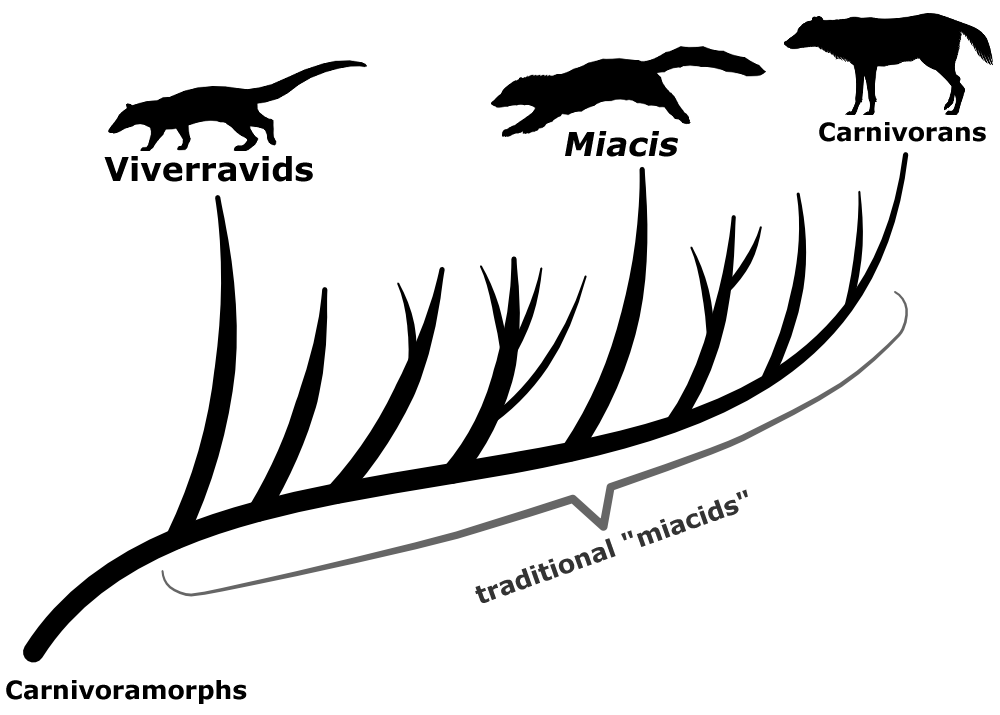
The earliest known carnivoran-like forms – known as carnivoramorphs – all looked vaguely-genet-like and were an ecologically diverse bunch of small predators, ranging from weasel-sized tree-climbers to fox-sized ground-based hunters, found all across North America and Eurasia during the Paleocene and Eocene. They lacked most of the anatomical specializations of true carnivorans, and didn’t quite fit into either the feliforms or caniforms, but their distinctive carnassial teeth make it obvious they were still very closely related.
From their initial discovery in the late 19th century, through to the late 20th century, these carnivoramorphs were traditionally all lumped together under the name “miacids“. As a result the group quickly turned into a big wastebasket taxon of similar-looking animals, all united more by just not being true carnivorans than by any shared characteristics between themselves.
Miacis parvivorus
But during the last couple of decades this mess has finally started to get cleared up. One distinct lineage of miacid-like animals called viverravids were split off, now thought to be the one of very earliest branches of the carnivoramorph evolutionary tree. Several other “miacids” have also been reassessed and renamed, reclassified as falling into various points in an evolutionary grade between viverravids and true carnivorans, and a coupleof species even turned out to actually be caniforms.
The true carnivorans arose from somewhere within the “miacids” during the mid-Eocene, but it’s still unclear where exactly to draw the taxonomic line between them. Forms like Quercygale and Tapocyon might be very close to the ancestral carnivoran – but they might instead be early feliforms – and some studies have also proposed that nimravids (“false sabertooth cats”) may actually be “advanced” carnivoramorphs instead of early feliforms.
There are also quite a few remaining “miacids” that still need sorting out, especially in the genus Miacis. There have to be other distinct lineages of these carnivoramorphs still hidden in the remaining wastebasket pile, and if we can eventually distinguish them from each other it might help to make early carnivoran relationships a bit clearer.
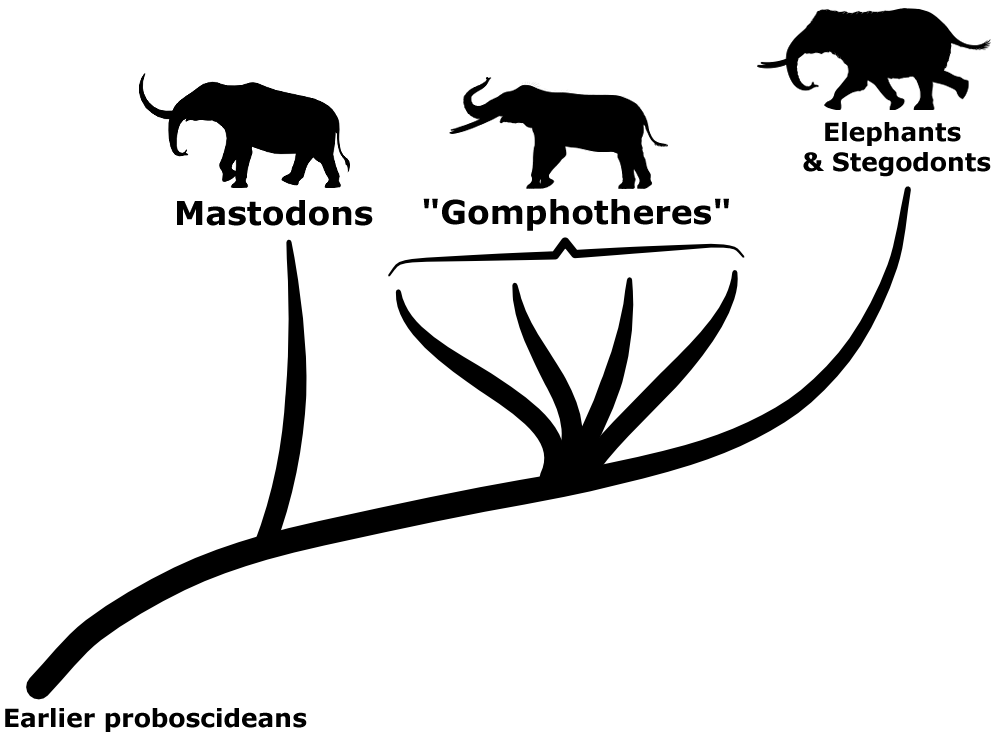
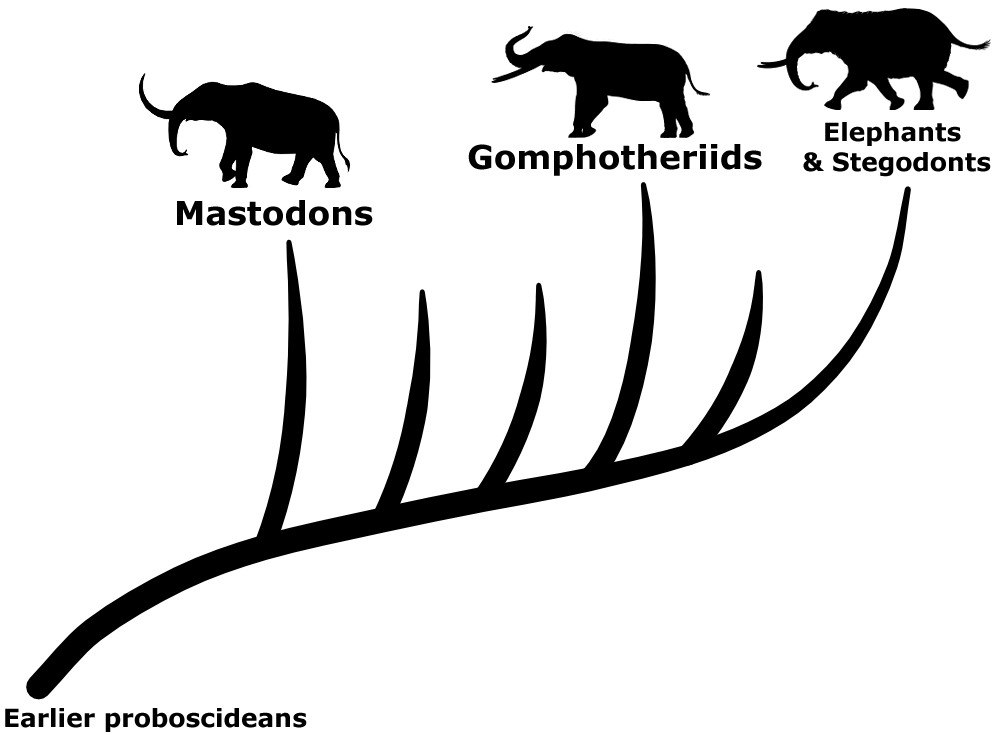
The three living species of elephants are the last surviving members of the proboscidean lineage – but up until the end of the last ice age about 11,000 years ago their relatives were much more numerous and widespread, found on every continent except Australia and Antarctica. Mammoths are probably the most famous of these recently-extinct proboscideans, closely related to modern Asian elephants, but there were also the more distantly-related stegodonts and mastodons…
Traditionally any proboscideans that fell into the evolutionary grade between mastodons and elephants-and-stegodonts were all labelled as gomphotheres. As a result by the late 20th century this group ended up as a wastebasket full of elephant-like forms that didn’t easily fit anywhere else, defined more by what they weren’t rather than by any features they all had in common.
This big collection of gomphotheres was highly diverse. Some species independently evolved similar convergent features, and there was also considerable individual physical variation within species, making the actual taxonomy of these animals very difficult to figure out. But over the last few decades there’s been a lot of revision of proboscidean evolutionary relationships, and gradually the gomphothere wastebasket has been clearing up. Groups like the choerolophodontids, amebelodontids, and anancids have been split off, leaving a more defined lineage of gomphotheres that do have shared anatomical characteristics – distinctive three-lobed trefoil-shaped wear patterns on their molar teeth.
These gomphotheriids were widespread, found across Africa, Europe, Asia, and the Americas – and they were especially successful in the latter. They arrived in North America during the Miocene (~16 million years ago) via the Beringia land bridge, and rapidly spread across the continent and down into Central America. They went on to become the only proboscideans to disperse into South America during the Great American Biotic Interchange, with two different lineages arriving at separate times – Notiomastodon around 2.5 million years ago, and Cuvieronius around 750,000 years ago.
Cuvieronius hyodon, a 2.3m tall (7’7″) South American gomphotheriid with distinctive spiraled tusks.
The exact relationships of the gomphotheriids to other elephant-like proboscideans are still a little uncertain. Both protein sequences and mitochondrial DNA have recently been recovered from 35,000-13,000-year old Notiomastodon specimens, but these studies have given different taxonomic conclusions – with the protein results suggesting gomphotheriids were most closely related to mastodons, and the DNA results suggesting they were much closer to true elephants.







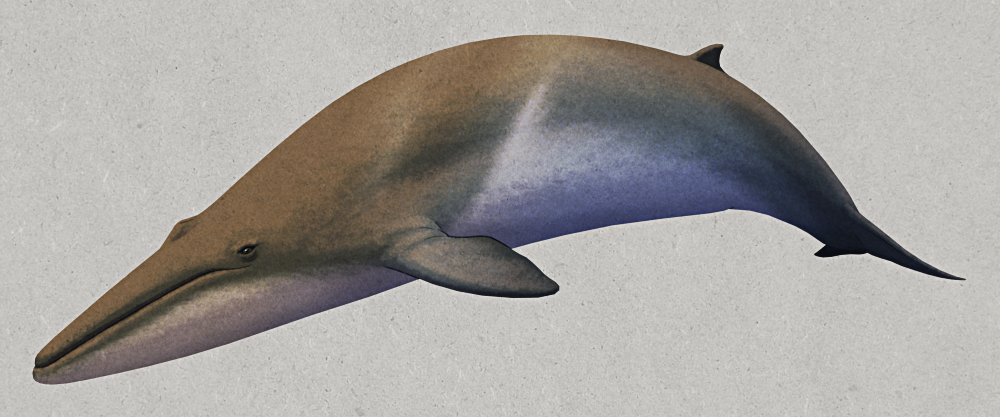




{kind=link}
{kind=link}
{kind=link}