

Isla Pedro González is part of the Pearl Islands in the Gulf of Panama, about 48km (30 miles) offshore. It was formed after the end of the last glacial period, as steadily rising sea levels cut it off completely from mainland Panama about 8500 years ago — and isolating the population of deer that lived there.
Although Pedro González is the third largest of the Pearl Islands it’s still fairly tiny, just 6.5km long (4 miles). As a result of this small space, limited available food supplies, and a lack of any large predators, these deer rapidly shrank down into a dwarfed form to survive. By 6000 years ago they were the size of a small dog, just 35-40cm tall at the shoulder (1’2″-1’4″) and weighing less than 10kg (22lbs).
The Pedro González dwarf deer haven’t been given any official scientific name just yet, but since they’re thought to be descendants of brocket deer they’d be a part of the genus Mazama, either as their own separate species or as a subspecies — similar to the larger native deer on nearby Isla San José, which are probably their closest living relatives.
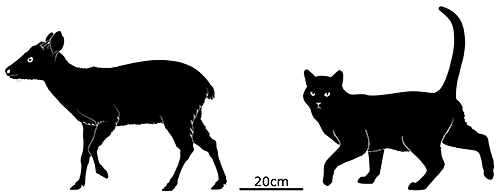
Paleoindian settlers arrived on Pedro González just over 6000 years ago, and we know they hunted and ate the tiny deer because all the known remains come from a preserved trash heap and show signs of human butchering and chewing.
Younger deposits have gradually less and less deer bones, and although those particular settlers had left again by about 5500 years ago the damage was already done — layers from another group of people about 2300 years ago show no deer bones at all, so the dwarf deer had to be already extinct by that time.













{kind=link}