

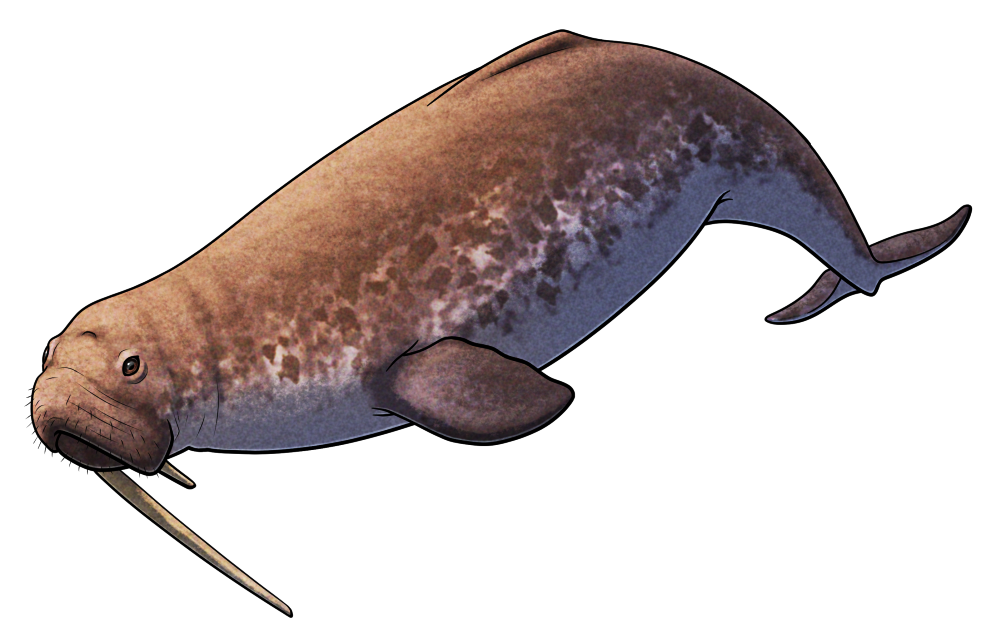
Maghriboselache mohamezanei was a cartilaginous fish from the late Devonian Period, about 369 million years ago, living in the shallow marine waters that covered what is now the Anti-Atlas mountain range of Morocco in northwest Africa.
Up to around 2.5m long (~8′), it’s known from several exceptionally well-preserved and near-complete skeletons.
It had a streamlined body with large pectoral fins, small pelvic fins, and a strongly keeled crescent-shaped tail fin. And although it was superficially shark-like in appearance, it was actually part of a lineage known as cladoselachids, which were much closer related to modern chimaeras than to sharks.
It’s unclear if Maghriboselache had two dorsal fins like its close relative Cladoselache, but some specimens preserve evidence of a chunky spine where the front dorsal fin would have been. Others show no sign of a front dorsal fin or spine at all, suggesting there may have been some sexual dimorphism going on in this species, with males having a spine (and possibly also an associated front dorsal fin) and females only having a rear dorsal fin.
But the most unusual feature of Maghriboselache was its nose.
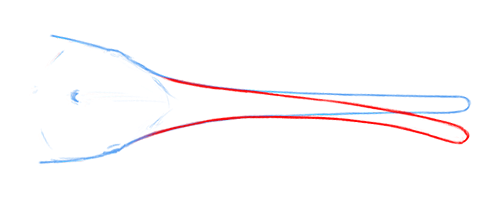
It had a very broad snout with large and unusually widely-spaced nostrils, which would have given it the ability to “smell in stereo” and determine the direction of scents carried through the water much more precisely – making it the earliest known example of that sort of sensory specialization.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}