Ergilornis rapidus, a 1.2-1.5m tall bird (4′-5′) from the Early Oligocene of Mongolia (~33-28 mya). Closely related to modern cranes, trumpeters, and limpkins, it was part of an extinct group called eogruids – flightless birds which existed across Eurasia for a large portion of the Cenozoic from roughly 40-3 million years ago.
Although the earliest known eogruids were smaller and less specialized, and may even have still been somewhat capable of flying, later forms like Ergilornis had highly reduced wings, long legs adapted for running, and convergentlyostrich-like feet with only two toes each.
Known from several near-complete fossils that include rare soft tissue impressions, it’s the first varanopid to show preserved skin details – revealing a pattern of very lizard-like rectangular scales. If it is a synapsid this is a pretty big deal, since early synapsids were previously thought to have had scale-less leathery skin.
It also had unusual mosaic-like patches of tiny osteoderms above its eyes, a feature previously known only in some temnospondyl amphibians. Whether this was the result of convergent evolution or the trait actually being ancestral to most tetrapods is unclear.
Its slender body, long digits, and highly curved claws indicate it was an agile climber. It probably mainly lived up in the treetops, feeding on insects, making it one of the earliest known tetrapods specialized for an arboreal lifestyle.
(*Maybe. There’s apparently an upcoming study that suggests varanopids might actually be sauropsids instead.)
Its short toothless snout suggests it was a suction feeder, using water pressure differences to pull small soft-bodied prey straight into its mouth like a syringe. Along with a heavily built body similar to those of hupehsuchians, and a very long tail that made up over half of its 1.6m length (5′3″), it was probably a fairly slow swimmer living in shallow coastal waters.
It was a close relative of Cartorhynchus, and may have been similarly capable of hauling itself onto land like a modern pinniped.
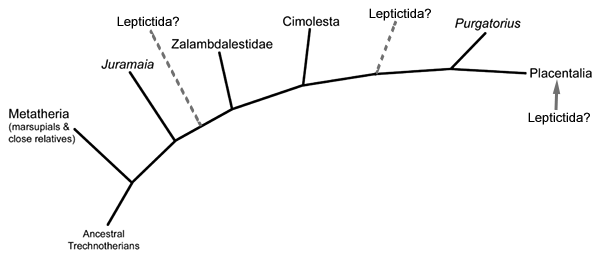
We don’t know exactly when placentals originated. The first definitive fossils come from the start of the Cenozoic, but a few early ancestral forms probably already existed during the Late Cretaceous (estimated up to 90-75 mya) and only got their chance to rapidly diversify immediately after the mass extinction event.
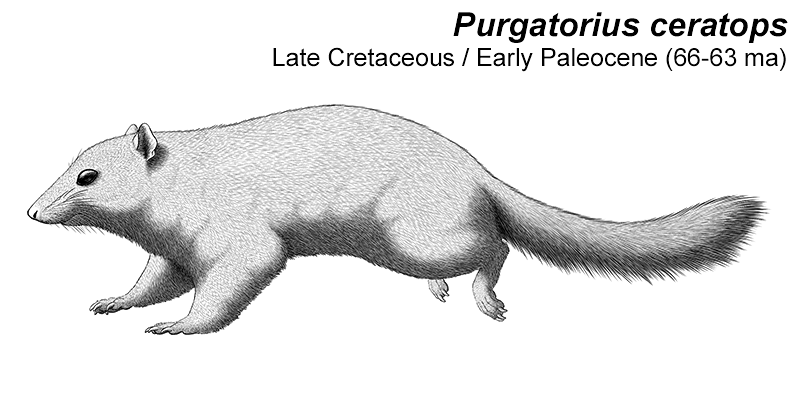
One of the closest fossils we have to the earliest placentals is Purgatorius. Known mainly from teeth from the Early Paleocene of North America (66-63 mya), it’s not entirely clear whether it actually existed in the Mesozoic, but its remains have been found very close to the K-Pg boundary and one fossil might actually be from the end-Cretaceous.
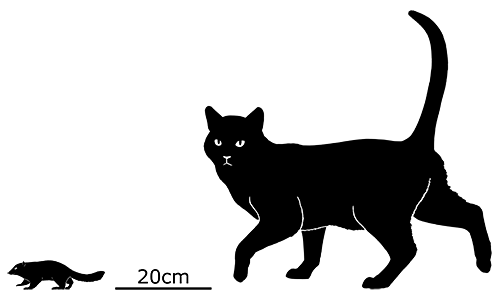
A few foot bones have been associated with some of the fossil teeth, and if they do belong to Purgatorius then they show that it had very flexible ankles, a characteristic typical of tree-climbing animals. It would likely have been a squirrel-like creature, about 15-20cm long (6-8″), eating an omnivorous mixture of insects, seeds, and fruit. It may also have been capable of burrowing similar to modern chipmunks.
It’s often been interpreted as a placental mammal, specifically a very early type of primate, but more recent studies suggest it might not even be a true placental at all – although it was probably still a very close relative of the common ancestor of all living placentals.
A group known as the leptictidans were probably some of the weirdest early eutherians. With their tiny forelegs, big hindlegs, and long counterbalancing tails, they somewhat resembled jerboas or small kangaroos – except they also had long slender snouts that probably ended in sengi-like proboscises, and their feet were structured more like those of running animals than jumping ones. They’re also thought to have been mainly bipedal, convergently evolving a similar posture and movement style to non-avian theropod dinosaurs.
First appearing in the Late Cretaceous, they made it through the end-Cretaceous extinction and survived up until the mid-Cenozoic across the northern hemisphere, going extinct around 33 million years ago. They were probably omnivores, eating a mixture of insects, small vertebrates, and soft plant matter such as fruit and leaves.
Their mix of “primitive” skull features and highly specialized skeletons makes classifying them particularly difficult. They’ve been proposed to be placentals related to primates and rodents or afrotheres, a very early branch of the eutherians, or close to placentals but not quite true members themselves. The latter interpretation currently seems most likely, but they could also be a paraphyletic group at the base of placentals (suggesting that they could even be ancestral to placentals, and therefore all placentals would technically be leptictidans).
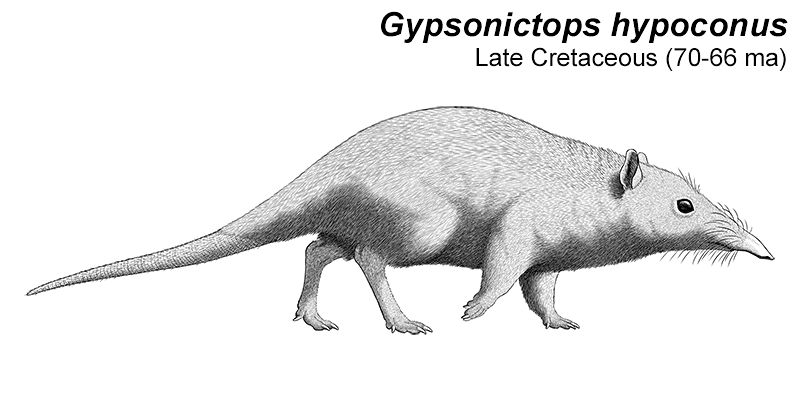
Gypsonictops was one of the earliest leptictidans, living during the Late Cretaceous of North America (70-66 mya). Known only from teeth and jaw fragments, we don’t know much about its appearance or full size – although it was probably smaller than its later relatives, perhaps about 35cm long (1′2″).
Any reconstruction of such fragmentary remains is going to be very speculative, but I’ve restored it here as a sort of transitional form, not yet quite as specialized. A more sengi-like animal, mainly quadrupedal but able to run and hop on its hind legs to flee from danger or chase after small fast-moving prey.
First appearing in the Late Cretaceous, a widespread and diverse group of mammals known as cimolestans were once thought to be early members of placental groups like pangolins and carnivorans. But more recent studies have shown them to be part of a different branch of the eutherian family tree altogether, more like cousins to the earliest placentals and leaving no living descendants.
One group of North American cimolestans, the taeniodonts, were specialized for digging up tough roots and tubers, with large claws, strong blunt jaws, and big front teeth that became ever-growing in some species.
Schowalteria was the earliest known member of this group, living during the Late Cretaceous of Canada (70-66 mya). Only represented by partial skull material, its full size is unknown – some estimates put it at a similar size to giants like Repenomamus, but it was likely closer to half that size at around 50cm in length (1′8″). Still one of the larger Mesozoic mammals around, but not nearly as big as some of the Cenozoic taeniodonts would later become.
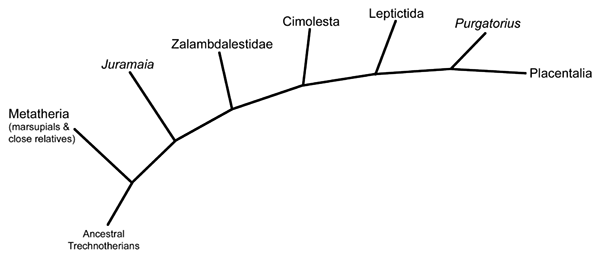
Living during the Late Cretaceous of Mongolia (85-70 mya), Zalambdalestes was part of a highly specialized group of mammals that it lends it name to – the zalambdalestids – which were an early branch of the eutherian evolutionary tree.
About 20-25cm in length (8-10″), it had relatively long limbs with especially strong hindlegs that show adaptations for rabbit-like hopping. Its long narrow snout may have ended in a flexible proboscis similar to those of modern sengi, and sharp interlocking teeth indicate a carnivorous or insectivorous diet.
Its long rodent-like incisors grew continuously throughout its life, suggesting it was gnawing on something tough enough to constantly wear down its front teeth.
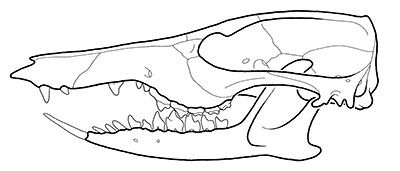
Skull of Zalambdalestes || from fig 51 in Wible JR, Novacek MJ, Rougier GW (2004) New data on the skull and dentition in the Mongolian late Cretaceous eutherian mammal Zalambdalestes. Bulletin Of The American Museum Of Natural History 281:1-144 uri: http://hdl.handle.net/2246/449
Some studies have proposed that zalambdalestids were actually very basal members of placental mammal groups such as rodents or rabbits, but the presence of epipubic bones in front of their pelvises (bones not found in placentals) shows they were a much earlier type of eutherian that still reproduced more like marsupials. Any anatomical similarities to later placentals were probably just the result of convergent evolution.
The other major branch of the therian mammals are the eutherians – represented today by only the placentals. But although the first eutherians split from their common ancestor with the metatherians all the way back in the mid-Jurassic, more then 160 million years ago, true placentals don’t seem to have actually appeared until right after the end-Cretaceous mass extinction.
There were many other side-branches of the eutherian lineage during the Mesozoic, however, and these last few days of the month will cover some of them.
(Some of these relationships are still under dispute, such as the exact position of leptictidans and Purgatorius and whether they’re actually placentals or not. Eutherian phylogeny is a bit of a mess.)
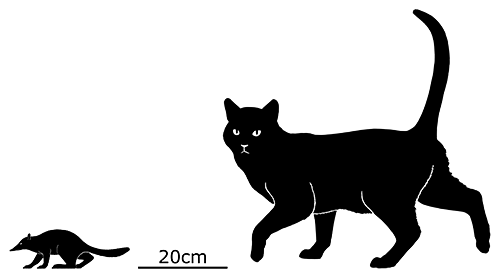
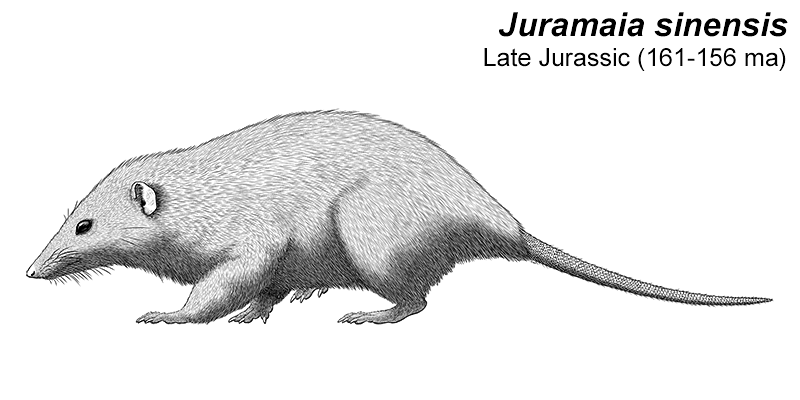
Juramaia was the earliest known eutherian, living during the Late Jurassic of China (161-156 mya). About 15cm long (6″), it was a shrew-like insectivore with limb anatomy that would have allowed it to climb up trees in a similar manner to modern rats.
Something very similar to it would have been the common ancestor of all later eutherians – suggesting that the earliest members of the group may have started out as tree-climbers before diversifying into different niches later on.
But despite it being closer related to living placentals than to marsupials, placental-style reproduction hadn’t actually evolved yet and it would have still given birth to tiny undeveloped young.
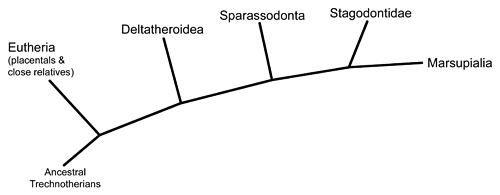
While true marsupials didn’t appear in the fossil record until the early Cenozoic, some of their closest relatives in the Mesozoic were a group called stagodontids. These metatherians evolved in North America during the Late Cretaceous, and much like the deltatheroideans seem to have taken over some of the ecological niches left vacant after the extinction of most eutriconodonts.
Their strong jaws and large blunt premolar teeth were adapted for crushing hard-shelled food, and they seem to have been semi-aquatic swimming animals (somewhat similar to modern water opossums) specialized for eating freshwater invertebrates such as snails and crustaceans.
Most stagodontids went extinct during the end-Cretaceous mass extinction, but if the early Cenozoic genus Eobrasilia is a member of the group then at least some survived in South America up until about 52 million years ago.
Other metatherians persisted in Europe, Africa, and Asia for most of the Cenozoic, with some fossils dating to just 11 million years ago, and the sparassodonts were successful in South America until about 3 million years ago – but today the only living members of this branch of the therian lineage are the marsupials in the Americas and Australasia.
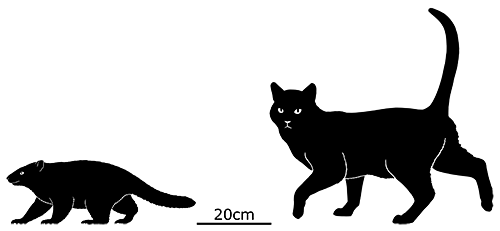
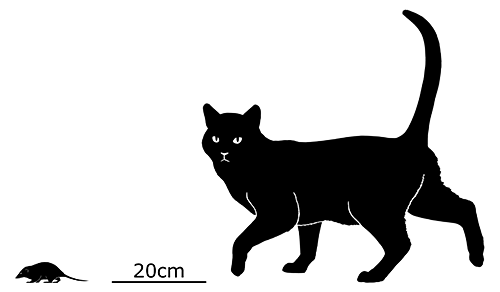
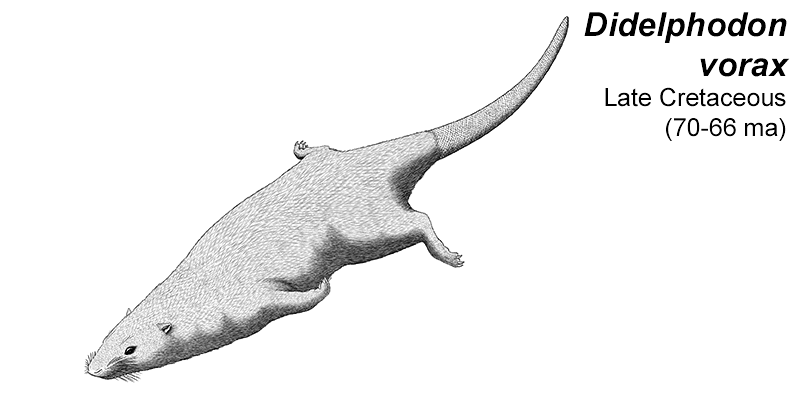
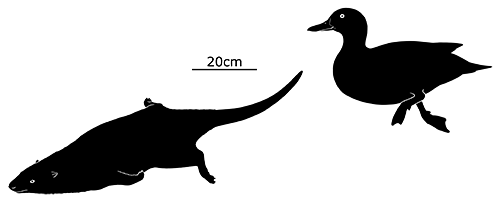
Didelphodon was a stagodontid living during the Late Cretaceous of North America (70-66 mya), and is known from skull and jaw remains, along with a fairly complete skeleton that hasn’t been officially described yet.
It had a long otter-like body and flexible feet, adaptations for efficient swimming, and was one of the largest known mammals at the time, measuring around 1m in length (3′3″).
Its sturdy jaws had an especially powerful bite force, one of the strongest relative to body size known for any mammal, and some of its molars were blade-like and similar in appearance to carnassials. Along with crunching on shellfish it would also have been capable of crushing bones and tough plants, and its diet was probably an omnivorous mixture of invertebrates, vegetation, carrion, and small vertebrates – potentially including other mammals and small dinosaurs.
In modern times the therian mammals are represented by just two surviving groups – placentals and marsupials. But both of these lineages contained many other extinct close relatives, and this final week of the month will focus on a few of them.
Modern marsupials are part of a larger grouping known as metatherians which split off from their common ancestor with placentals during the Jurassic period, at least 160 million years ago. Probably originating in Asia, they spread to Europe and the Americas during the Cretaceous, and diversified into several different groups – including some which would go on to become the dominant mammalian predators in South America during its long period of isolation in the Cenozoic.
One of the earliest branches of the metatherians were the deltatheroideans. These mammals are known from Asia and North America, evolving to fill vacant small carnivore niches after the disappearance of most of the eutriconodonts. Most of them went extinct at the end of the Cretaceous, but a few did survive for at least 10 million years afterwards.
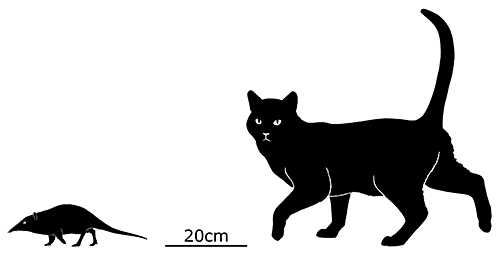
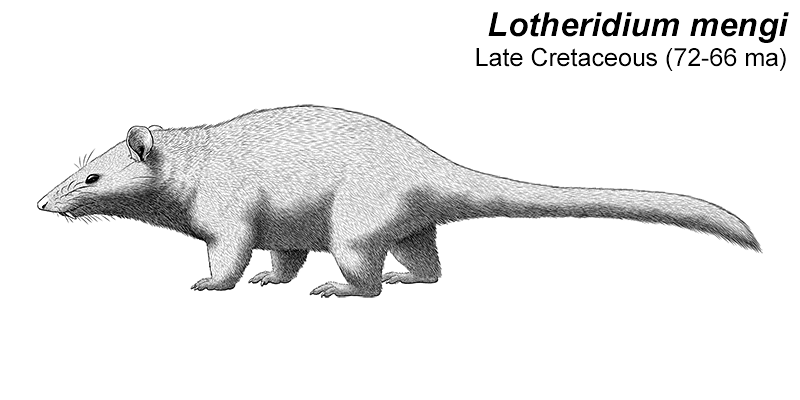
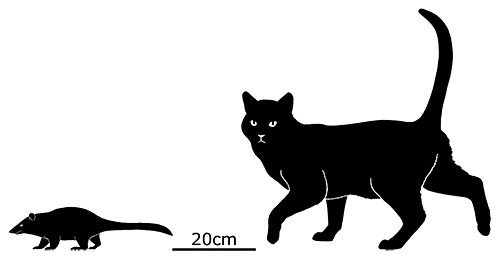
Lotheridium was a deltatheroidean living during the Late Cretaceous of China (72-66 mya), and is known from a nearly complete skull. It’s likely to have had a head-and-body length of about 20cm (8″), and may have had a full length including the tail of up to 30-40cm (1′-1′4″).
It had elongated canine teeth, convergently similar to the saberteeth of manylatermammalgroups. It was probably a highly specialized predator, and may even have been capable of preying on small dinosaurs – its close relative Deltatheridium is known to have eaten theropods like Archaeornithoides.







{kind=link}