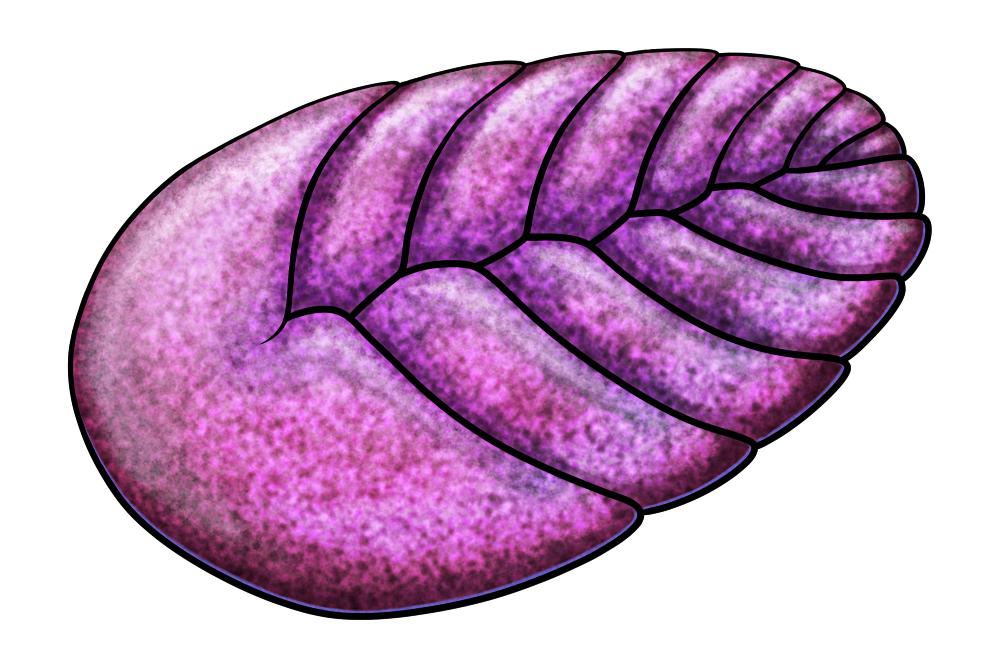
It was a small worm-like animal about 1.6cm long (~0.6″), with bundles of long bristles along its sides and flap-like structures on its underside. Its back was also covered with sclerite armor arranged in a strangely asymmetrical fashion, with larger overlapping plates in the middle and numerous smaller cap-like sclerites distributed unevenly along each side.
Its discovery actually confirms an old prediction that lophophorates probably originated from armored worm-like animals, representing an evolutionary link between earlier free-living annelid-like forms and later immobile filter-feeding tommotiids.
It’s not known why the armor on Wufengella‘s back was so unevenly organized – but some of the later tube-like tommotiids also had weird symmetry going on, with forms like Eccentrotheca having irregular sclerites arranged in a spiral around their bodies.
Most animals are bilaterally symmetric, having body plans with mirrored left and right sides – which also allows them to have a defined head end, rear end, top side, and underside.
It’s not entirely clear what evolutionary advantage this type of symmetry gave to the first bilaterians, which would have been been small “simple” worm-like animals living sometime during the Ediacaran Period between 600 and 560 million years ago. The current generally accepted explanation is that it probably allowed for better active locomotion – clustering sense organs at the head end and directing body movement more efficiently towards food sources and away from threats.
…But not every bilaterian has stayed roughly symmetrical.
Over the last half-billion years or so some bilaterians have abandoned their roughly-mirror-image body plans in favor of something distinctly wonkier. Asymmetry has evolved multiple times in various different lineages, and so every weekday this month we’ll be looking at some examples.
And we might as well start way back near the beginning:
Strange Symmetries #01: Almost Bilateral
Living in the Ediacaran between about 567 and 550 million years ago, the proarticulatans were flattened rounded organisms with two rows of soft “quilted” rib-like segments (known as isomers) and sometimes a larger fused “head” section at the front. The left and right isomers weren’t perfectly mirrored, instead being offset from each other in a glide reflection pattern – but the presence of a clear central body axis suggests these animals may have had some sort of relation to the earliest bilaterians, possibly even being a very early stem group that was experimenting with a not-quite-totally-bilateral body plan.
Vendia sokolovi
Discovered in what is now northwest Russia, and dating to around 555 million years ago, Vendia sokolovi was a small proarticulatan measuring about 1.1cm long (0.4″). It had a rather small number of isomers compared to some of its relatives, only 7 per side, and seems to have had a simple digestive tract that branched into each isomer.
(The superficial resemblance to trilobites was coincidental – while we might not be entirely sure what these things were, we do at least know they weren’t closely related to early arthropods.)
Very little overall is known about these animals’ lifestyles. Trace fossils suggest they were able to move around, feeding on microbial mats on the seafloor, and they may also have been able to firmly stick themselves onto the spots they were currently grazing.
Lobopodians were some of the earliest known panarthropods, closely related to velvet worms, tardigrades, and the ancestors of all the true arthropods. They were small soft-bodied worm-like animals with multiple pairs of fleshy legs, and some species also bore elaborate spikes, armor plates, and fleshy bumps all over their bodies – with the spiny Hallucigenia being the most famous example.
But unlike its more charismatic relative Paucipodia inermis here didn’t seem to have any ornamentation at all.
Known from the Chinese Chengjiang fossil deposits, dating to about 518 million years ago, Paucipodia lived in what was then a shallow tropical sea. Its 13cm long (~5″) tubular body had nine pairs of legs, with each foot tipped with a pair of hooked claws, and the inside of its mouth was ringed with tiny sharp teeth.
Several specimens have been found preserved in association with the weird gummy-disc animal Eldonia, which may indicate Paucipodia either preyed on them or scavenged on their carcasses.
Some Paucipodia fossils also have enigmatic tiny “cup-like” organisms attached to their legs. It’s currently unknown what exactly these were, or whether they were parasitic in nature or simply opportunistically “hitching a ride” similar to the Inquicus found on armored palaeoscolecid worms in the same fossil beds.
It had a wingspan of up to 4m (~13′), with a head-and-body length of around 1.2m (~4′), and like other dsungaripterids it had a distinctively reinforced skull, with a toothless beak at the front of its jaws and strong widely-spaced teeth further back – an arrangement that was probably used to catch and then powerfully crack open hard-shelled prey.
Unlike other pterosaurs, however, Noripterus also had some very unusual feet.
All other known pterosaurs seem to have had plantigrade hindlimbs, standing and walking with the whole foot on the ground. But Noripterus had toe joints that looked more like those of theropod dinosaurs than other pterosaurs, with a higher level of upward flexibility and potentially a more digitigrade posture standing on just its toes.
With dsungaripterids already having fairly stout body proportions that suggest they spent a lot of time walking around on the ground, Noripterus may have been even more agile and adept at terrestrial locomotion. Digitigrady is generally more efficient for moving at higher speeds, so this pterosaur might have been a runner behaving similarly to modern ground birds, preferring to sprint away from threats on foot rather than launch itself into the air.
Known only from South America between about 242 and 205 million years ago, these reptiles’ heads were wide at the back but very narrow along the snout, often with prominent bony bumps and ridges on their skulls, and they had less osteoderm armor on their bodies than other archosauriformes.
They’ve traditionally been interpreted as very crocodile-like and semi-aquatic, but their long slender limbs and presence in rather arid paleoenvironments suggest they may have been more terrestrial fast-running predators.
Tropidosuchus romeri here lived about 235 million years ago in what is now Argentina. It was one of the smaller proterochampsids, only about 50cm long (1’8″), with just a single row of osteoderms along its back, and had larger and lower-set eyes compared to its relatives.
CT scans of its braincase indicate it had a particularly good sense of smell, and it may have relied mainly on scent to locate prey.
Antaecetus aithai was an early whale that lived during the late Eocene (~40 million years ago) in what is now Morocco, at a time when northern Africa was covered by a warm shallow sea.
It was part of the “basilosaurids“, some of the first fully aquatic cetaceans – traditionally considered to be a single defined group, but more recently found to be more of an “evolutionary grade” of multiple early whale lineages – and much like Basilosaurus it had elongated back vertebrae that would have given it a very long slender body shape.
Antaecetus also had a proportionally smaller head and smaller teeth than other basilosaurids, along with much denser bones and a stiffer spine that would have made it a rather slow swimmer with reduced maneuverability. It was also fairly small overall compared to most of its relatives, probably around 6m long (~20′).
It was probably a slow-moving coastal water animal somewhat like modern sirenians – except unlike manatees and dugongs it was carnivorous. Its relatively delicate teeth suggest it was feeding on soft-bodied prey like cephalodpods, and with its lack of speed it must have been some sort of ambush predator, waiting around for potential prey to come within striking range.
Towards the end of the Cretaceous, about 69 million years ago, the most diverse and numerous mammals in the northern hemisphere were the metatherians, close relatives of modern marsupials.
About the size of a modern mouse, around 10-15cm long (4-6″), and with teeth that suggest it was a shrew-like insectivore, this little metatherian lived in northern Alaska in what’s known as the Paaŋaqtat Province – a region with a distinctive population of endemic polar animals. At the time this area was located at an even higher latitude than it is today, around 80-85ºN, but due to a greenhouse climate it was also warmer, with no permanent ice and the average temperatures staying above freezing.
Unnuakomys was by far the most common mammal species in the Paaŋaqtat Province, represented by numerous fossil teeth and a few jaw fragments, and it also seems to have been the only metatherian living in the whole region. This may just be a preservation bias in the fossil record, but it might also indicate that Unnuakomys was uniquely specialized to endure the several months of continuous darkness each winter in its polar woodland environment, while other North American metatherians were restricted to more southerly latitudes.
Cabarzia trostheidei here lived during the early Permian in what is now Germany, about 295 million years ago.
Despite its very lizard-like appearance it was actually part of the varanopid lineage, a group of scaly amniotes traditionally classified as early synapsids (distant relatives of modern mammals), but which more recently have been proposed to instead be sauropsid reptiles closer related to early diapsids.
It was around 50cm long (1’8″), and its short arms, long legs, slender body, and long tail suggest it was capable of shifting into a bipedal posture when running at high speeds, similarly to some modern lizards – probably mainly to escape from larger predators, but possibly also used to pursue fast-moving prey like flying insects.
And whether varanopids were actually synapsids or sauropsids, this makes Cabarzia the earliest known example of an animal running on two legs.
Halszkaraptorines were a group of small dromaeosaurids known only from the Late Cretaceous of Mongolia. They were odd little raptors with flattened snouts, long necks, and flipper-like arms – features that suggest they were specialized for swimming, making them the second known lineage of semi-aquatic non-avian dinosaurs after the spinosaurids.
Natovenator polydontus lived in what is now the Gobi Desert in southern Mongolia, around 72 million years ago. The size of a small duck, about 45cm long (18″), it had jaws full of many needle-like teeth, a long flexible goose-like neck, and a streamlined body with a wide flattened ribcage convergently shaped like those of modern diving birds.
Although it had long strong legs, these don’t show much in the way of aquatic specializations and would have been used more for walking and running on land. Instead it may have used its flipper-like arms to propel itself through the water, like modern penguins or auks.
It probably had a lifestyle similar to modern mergansers, swimming and diving in lakes and rivers, and preying on fish, amphibians, and aquatic invertebrates.
Most of them also had jaws full of round flat teeth used to crush hard-shelled prey, but some may instead have been herbivorous grazers similar to parrotfish.
And a couple of lineages even became carnivores.
Serrasalmimus secans lived in what is now Morocco during the late Paleocene, about 59 million years ago. Although only known from its jaws, the size of the fossil material suggests it was fairly large for a pycnodont, possibly around 80cm long (~2’8″).
It had sharp flesh-cutting teeth similar to those of modern piranha, but with a surprising evolutionary twist. Unlike any other known ray-finned fish, Serrasalmimus‘ teeth were true shearing carnassials anchored into bony sockets, with new replacement teeth forming directly below each current tooth – a very specific arrangement of features previously only known in mammals.
This is an especially remarkable example of convergent evolution because on land placental carnivorans were developing their own carnassials at the same time, just a few million years after the K-Pg mass extinction. Both mammals and pycnodonts were simultaneously taking advantage of the vacant predatory roles in their respective ecosystems, and ended up with incredibly similar tooth adaptations as a result.











{kind=link}
{kind=link}
{kind=link}