

Hupehsuchians were small marine reptiles closely related to ichthyosaurs, known only from the Early Triassic of southwestern China about 249-247 million years ago. They had toothless snouts, streamlined bodies, paddle-like limbs, and long flattened tails, along with a unique pattern of armor along their backs made up of overlapping layers of bony osteoderms.
Hupehsuchus nanchangensis was a mid-sized member of the group, about 1m long (3’3″). Newly-discovered fossils of its skull show that its long flattened snout had a distinctive gap between the bones (similar to the platypus-like snout seen in its relative Eretmorhipis) with an overall shape surprisingly convergent with that of modern baleen whales – suggesting that this hupehsuchian may have been a similar sort of filter-feeder.
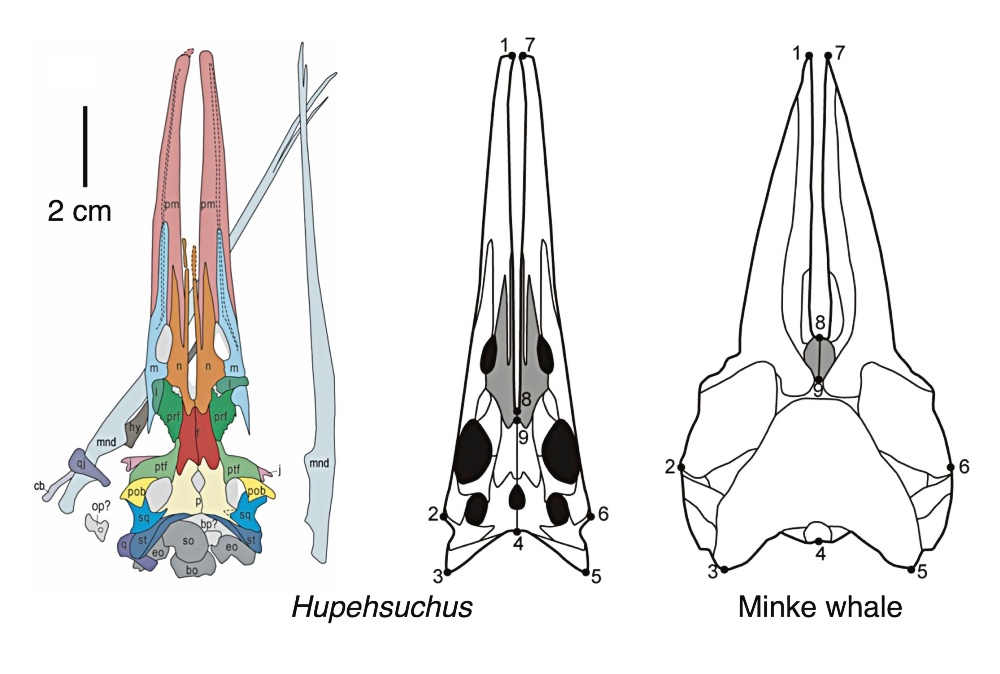
From fig 2 & fig 3 of Fang et al (2023). First filter feeding in the Early Triassic: cranial morphological convergence between Hupehsuchus and baleen whales. BMC Ecol Evo 23, 36. https://doi.org/10.1186/s12862-023-02143-9
Grooves in the bones along the outer edges of its upper jaws may be evidence of filtering structures similar to baleen, although with no soft-tissue preservation we don’t know exactly what this would have looked like. Its slender flexible lower jaws probably also supported a large expandable throat pouch, allowing it to filter plankton out of larger volumes of water.
















{kind=link}
{kind=link}